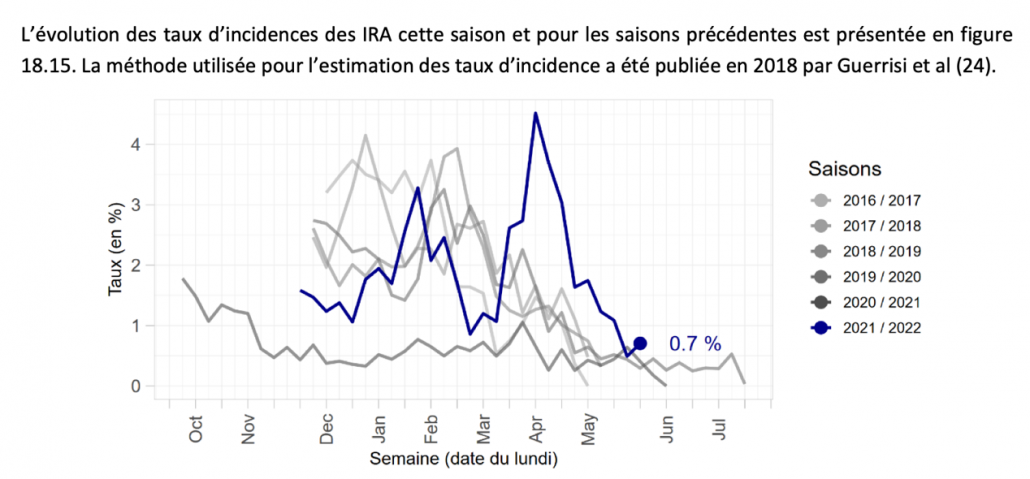
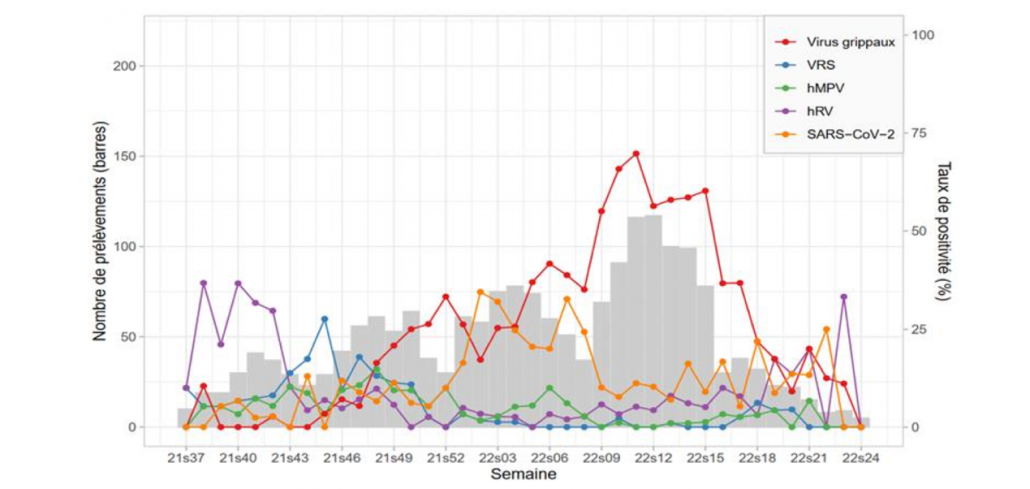
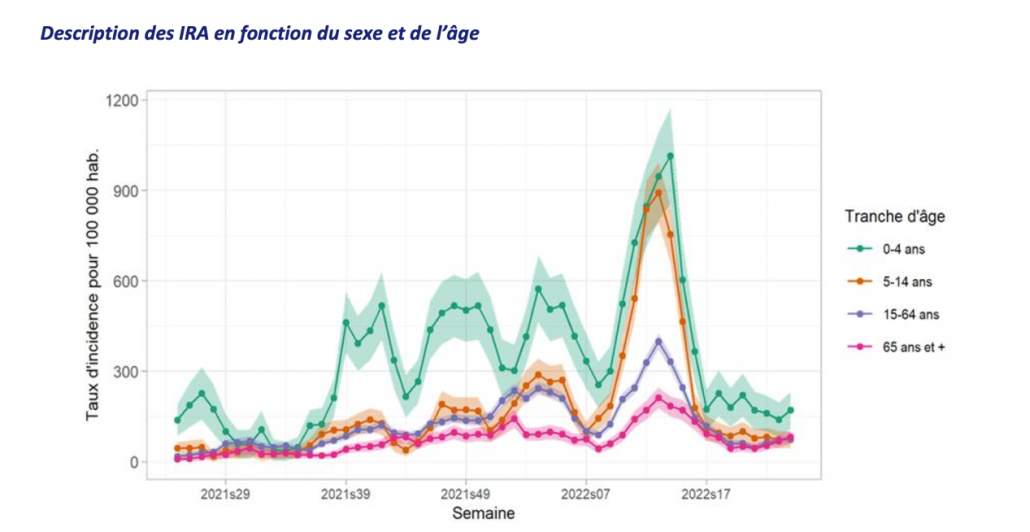
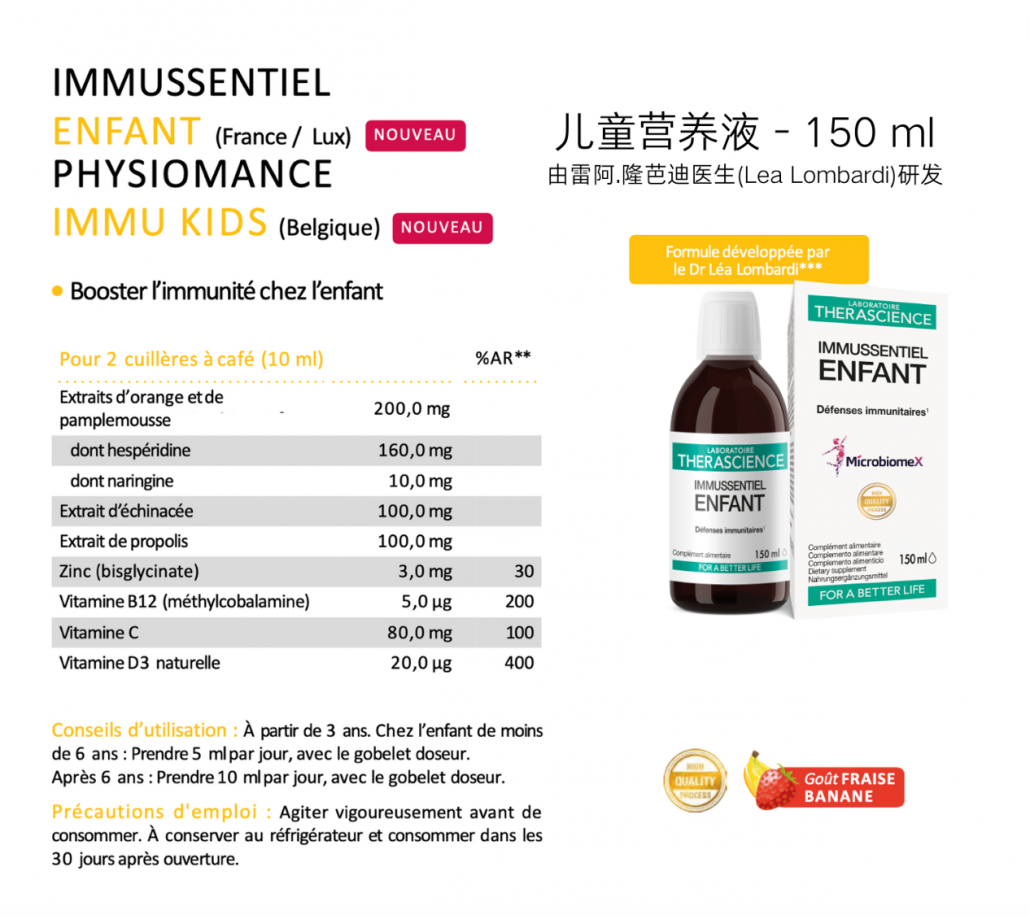
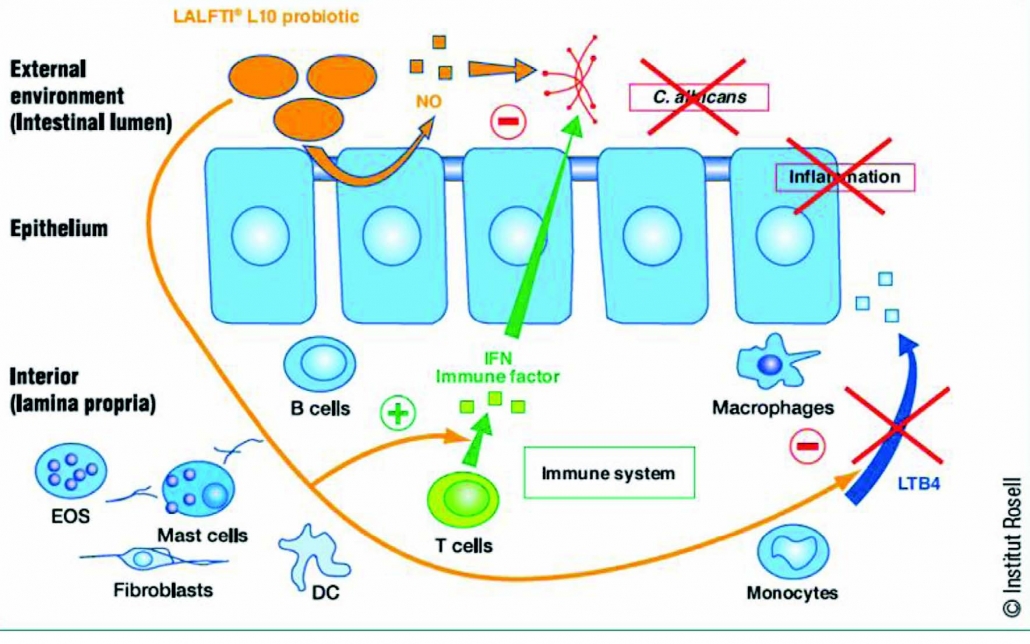
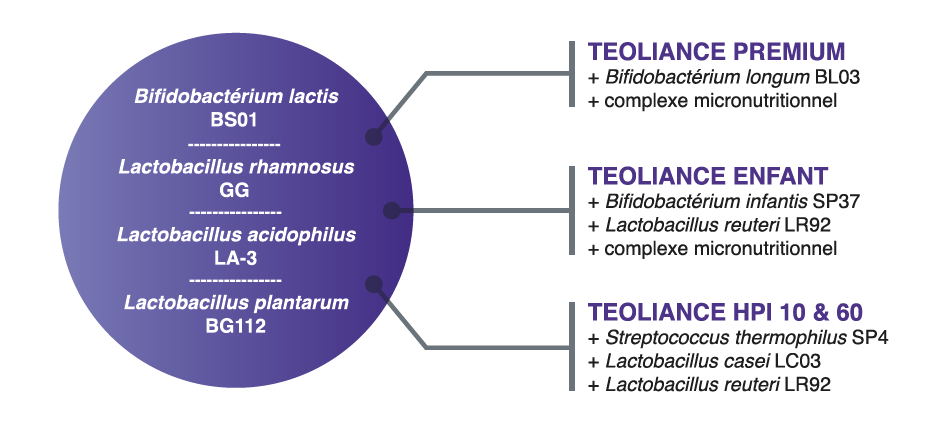
1.Postbiotics and Their Potential Applications in Early Life Nutrition and Beyond
2.Scavenger receptor for lipoteichoic acid is involved in the potent ability of Lactobacillus plantarum strain L-137 to stimulate production of interleukin-12p40.
3.Rational identification of diet-derived postbiotics for improving intestinal microbiota function
4.Probiotics for Gastrointestinal Conditions: A Summary of the Evidence
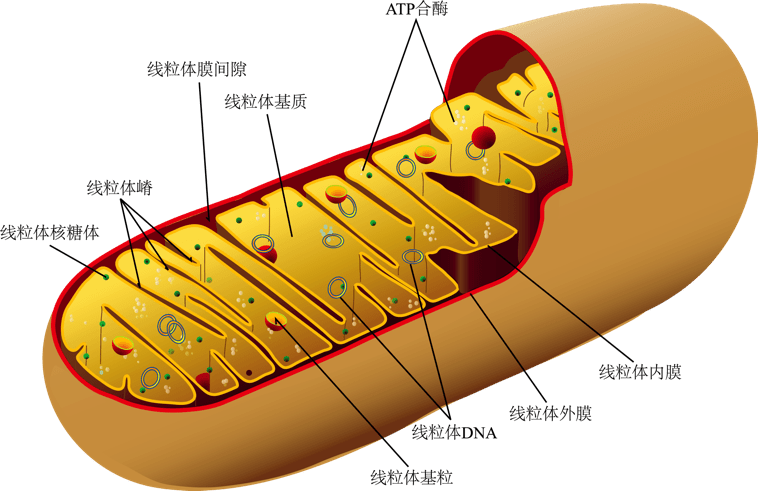
1.Nuo Sun, Richard J. Youle, Toren Finkel, Mitochondrial Theory of Aging and Other Aging Theories. Mol Cell. 2016 Mar 3; 61(5): 654–666衰老的线粒体基础
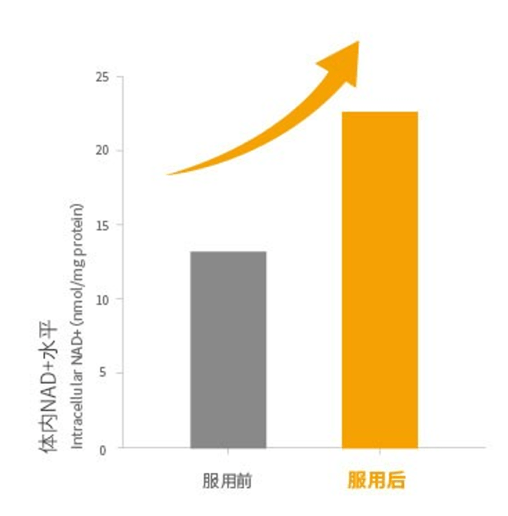
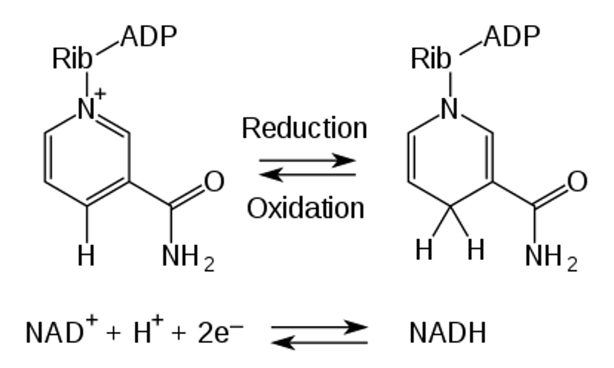
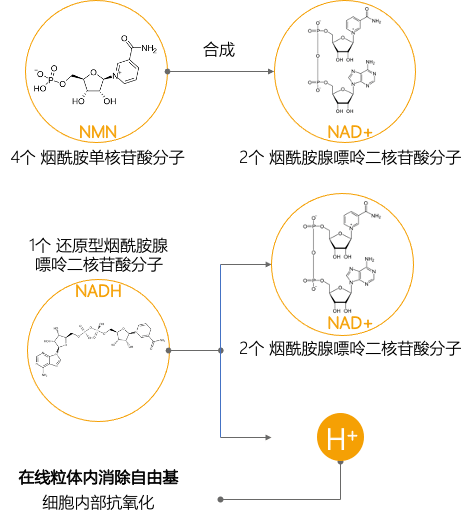
2.Birkmayer G. Nadh: The Energizing Coenzyme. McGraw Hill Professional, 1998 Nadh:增加活力的辅酶
3.Birkmayer. JGD. Coenzym-1 (NADH) – Eine wissenschaftlich bewiesene Anti-Aging-Substanz. Prevention and anti aging 2006: 340-348. 还原型辅酶I(NADH):经过科学证明的抗衰老物质
4.Nadlinger K, Westerthaler W, Storga-Tomic D, Birkmayer JGD. Extracellular metabolization of NADH by blood cells correlated with intracellular ATP levels. Biophys. Biochim Acta 2002; 1573: 177-182. 血细胞对NADH的细胞外代谢与细胞内ATP水平相关
5.Zhu K(1), Swanson RA, Ying W. NADH can enter into astrocytes and block poly(ADP-ribose) polymerase-1-mediated astrocyte death. Neuroreport. 2005 Aug 1;16(11):1209-12. NADH可以进入星形胶质细胞并阻断聚(ADP-核糖)聚合酶1介导的星形胶质细胞死亡
6.Demarin V, Podobnik-Sarkanji S, Storga-Tomic D, Kay G, Martinic-Popovic,M. Puretic B., Birkmayer JGD. ENADA/NADH improves cognitive impairment of Alzheimer patients. J. Tumor Marker Oncology 2003; 18:43-49. ENADA / NADH可改善阿尔茨海默氏症患者的认知障碍
7.Reibnegger G, Greilberger J, Juergens G. and Oettl K. The antioxidative capacity of ENADA®-NADH in humans. J. Tumor Marker Oncol. 2003; 18, 37-41. ENADA®-NADH在人体内的抗氧化能力
8.Lotfi R, Birkmayer J, Ghouni A, Metabolic and Anthropometric Effects of NADH RAPID Energy in Diabetic Type 2. Journal of Food Science and Engineering 6 (2016) 280-285. NADH RAPID能量在2型糖尿病患者中的代谢和人体测量作用
9.Birkmayer JGD, Vrecko C, Volc D, Birkmayer W.Nicotinamide adenine dinucleotide (NADH) – a new therapeutic approach to Parkinson’s disease: Comparison of oral and parenteral application. Acta Neurol Scand. 1993; 87: 32-35. 还原型烟酰胺腺嘌呤二核苷酸(NADH)-帕金森氏病的新治疗方法:口服和肠胃外应用的比较
10.Birkmayer JGD, Zhang JR, ed. Bagchi D, Preuss H. NADH in Cancer Prevention and Therapy. CRC Press 2005; chapter 33, 541- 554. NADH在癌症的预防和治疗中的应用
11.Grathwohl D, Klann M, Müller HM, Schlachter H, Berg A. Influence of NADH supplementation for the muscular energy supply in humans. Deutsche Zeitschrift für Sportmedizin, 11/2000. 补充NADH对人类肌肉能量供应的影响
12.Kay GG, Virre ES, Moline M. Stabilized NADH (ENADA) as Counter-Measure Jet Lag and Sleep Deprivation. J. Tumor Marker Oncol. 2001; 16 : 124-125. 稳定的NADH(ENADA)作为应对时差和睡眠不足的措施
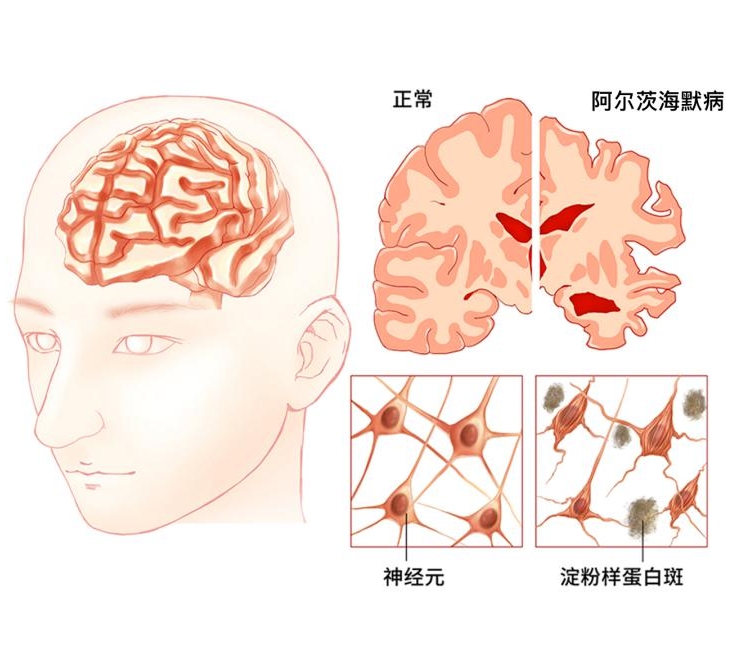
Association AS. 2015 alzheimer’s disease factsand figures [J]. Alzheimers & Dementia the Journal of the Alzheimers Association, 2015,11(3):332-384.
Jia J, Wang F, Wei C, et al. The prevalence of dementia in urban and rural areas of china [J]. Alzheimers & Dementia the Journal of the Alzheimers Association, 2014,10(1):1-9.
贾建平, 陈生弟. 神经病学.第7 版[M]. 人民卫生出版社, 2013.
Plataras C, Tsakiris S, Angelogianni P. Effect of cdp-choline on brain acetylcholinesterase and na(+), k(+)-atpase in adult rats [J]. Clinical Biochemistry, 2000,33(5):351-357.
Pinardi G, Pelissier T, Kramer V, et al. Effects of cdp-choline on acetylcholine-induced relaxation of the perfused carotid vascular beds of the rat [J]. General Pharmacology the Vascular System, 1994,25(4):635-638.
Petkov VD, Konstantinova ER, Petkov VV, et al. Learning and memory in rats exposed pre- and postnatally to alcohol. An attempt at pharmacological control [J]. Methods & Findings in Experimental & Clinical Pharmacology, 1991,13(1):43-50.
Secades JJ. Citicoline: Pharmacological and clinical review, 2016 update [J]. Revista De Neurologia, 2016,63(S03):S1.
Castagna A, Cotroneo AM, Ruotolo G, et al. The citirivad study: Citicoline plus rivastigmine in elderly patients affected with dementia study [J]. Clinical Drug Investigation, 2016,36(12):1-7.
Gareri P, Castagna A, Cotroneo AM, et al. The citicholinage study: Citicoline plus cholinesterase inhibitors inaged patients affected with alzheimer’s disease study [J]. Journal of Alzheimers Disease Jad, 2017,56(2):557.
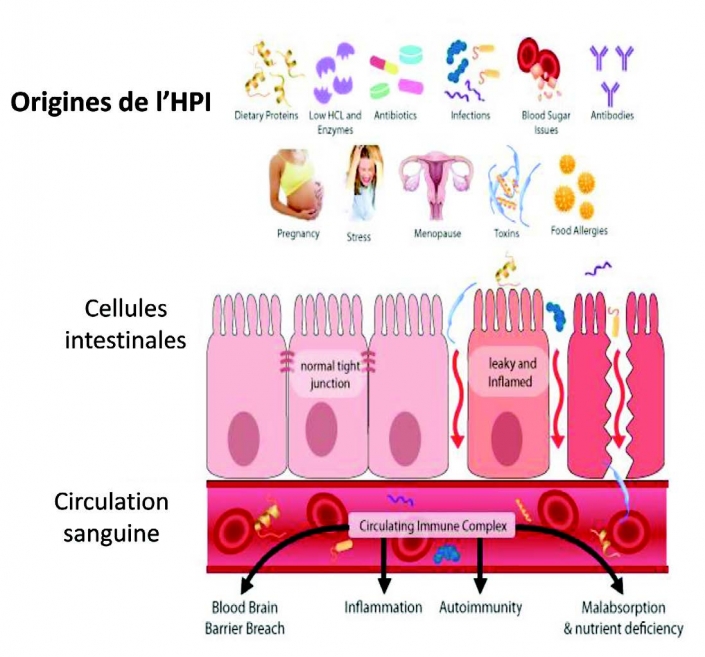
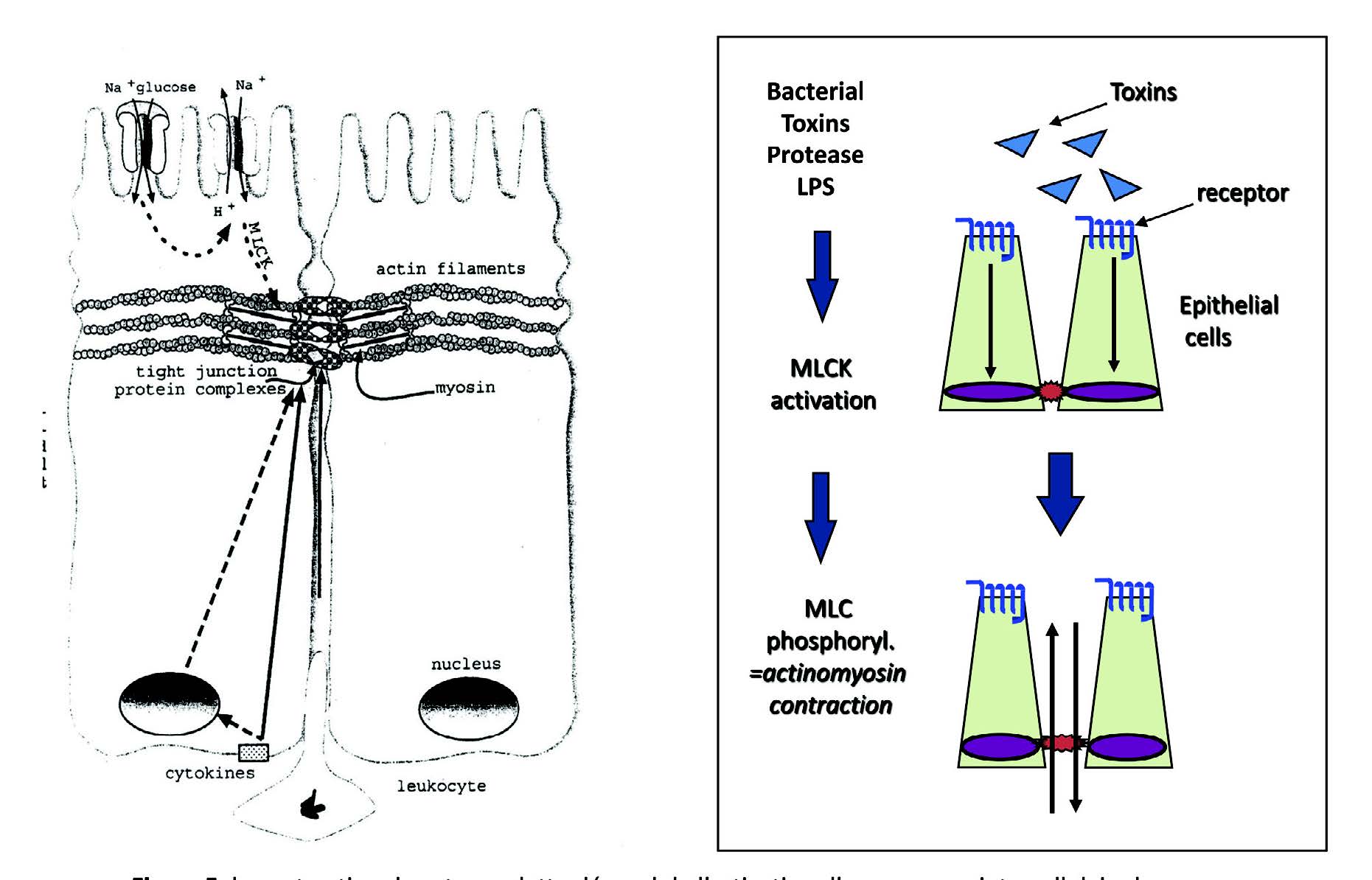
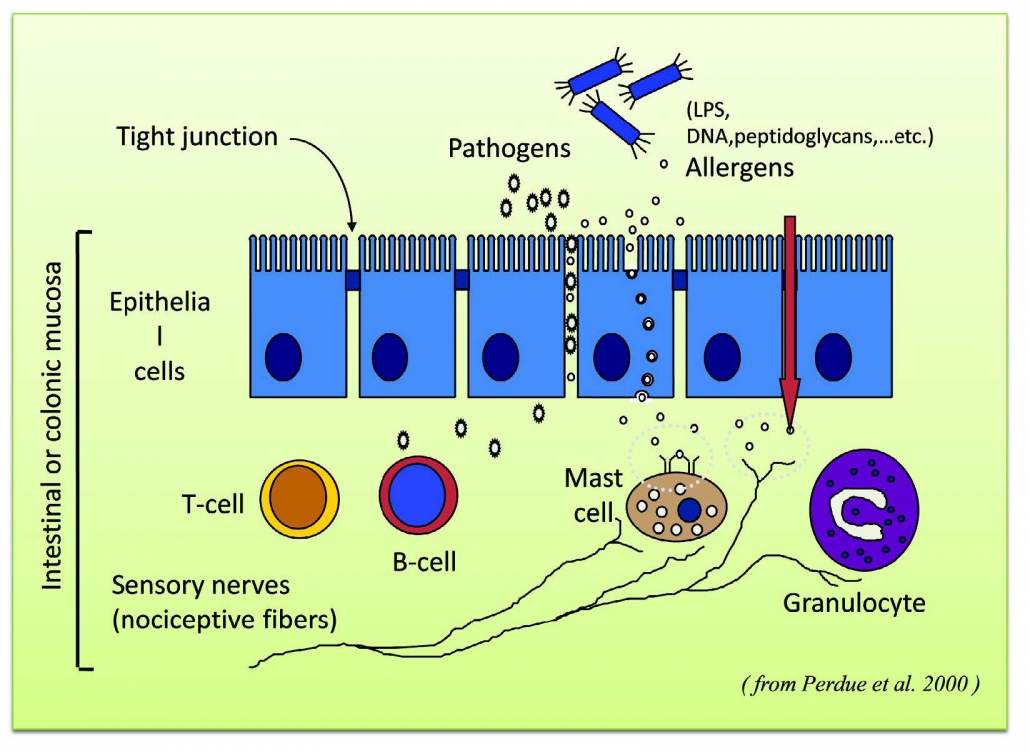
Visser J, Rozing J, Sapone A, Lammers K, Fasano Tight junctions, intestinal permeability, and autoimmunity: celliac disease and type I diabetes paradigms. 紧密连接,肠道通透性和自身免疫性:乳糜泻和I型糖尿病范例Ann N Y Azad Sci. 2009;1165 :195-205.
Annahazi A, Ferrier L, Bezirard V, Leveque M, Eutamema. H, Ait-Belgrooui A, Coeffier M, Ducrotte P, Roka R, Inczefi O, Gecse K, Rosztoczy A, Molnar T, Ringel- Kulka T, Ringel Y, Piche T, Theodorou V. Wittmann T, Bueno Luminal cysteine-porteases degrade colonic tight junction structure and are responsible for abdominal pain in constipotion-predominant IBS. 发光的半胱氨酸蛋白酶降解结肠紧密连接结构,并导致便秘为主的IBS腹痛Am J Gastroenterol 2013; 108 :1322-3.
Puthenedom M, Williams PH, Lakshmi BS, Balakrishnan. Modulation of tight junction barrier function by outer membrane proteins of enteropathogenic Escherichia coli: Role of F‐actin and junctional adhesion molecule‐1. 肠致病性大肠杆菌的外膜蛋白对紧密连接屏障功能的调节:F-肌动蛋白和连接黏附分子-1的作用Cell Biol 2007; 31: 836-44.
Nusrat A, von Eichel-Streiber C, Turner JR, Verkade P, Madara JL, Parkos CA. Clostridium difficile toxins disrupt epithelial barrier function by altering membrane microdomain localization of tight junction proteins. 艰难梭菌毒素通过改变紧密连接蛋白的膜微区定位来破坏上皮屏障功能Infect Immun. 2001; 69: 1329-36.
Shen L, Su L, Turner Mechanisms and functional implications of intestinal barrier defects. 机制和肠屏障缺陷功能的影响Dig Dis. 2009;27: 443-9.
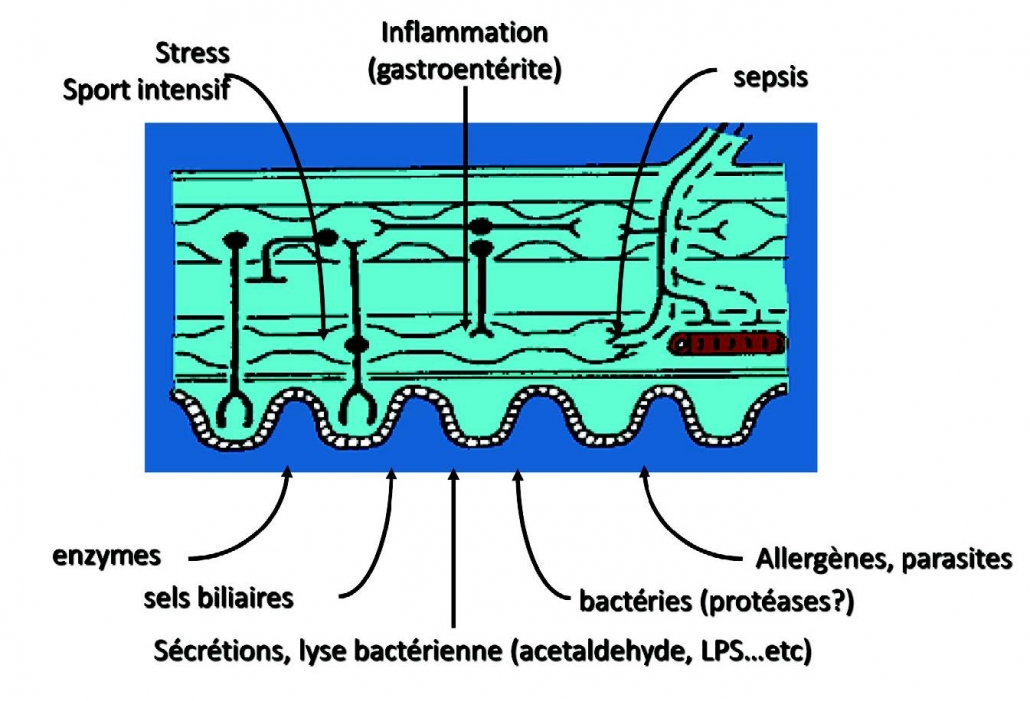
Da Silva S, Robbe-Masselot Ait-BeIgnaoui A, Mancuso A, Mercade-Loubiere M, Salvador Cartier C, Gillet Ferrier K, Loubiere R, Dague E, Theodorou V, Mercier-Bonin M. Stress disrupts intestinal mucus barrier in rats via mucin O-glycosylation shift prevention by a probiotic treatment. 应激通过益生菌治疗预防粘蛋白O-糖基化转移来破坏大鼠肠粘液屏Am J Physiol Gastrointest liver Physiol. 2014; 307: G420-9.
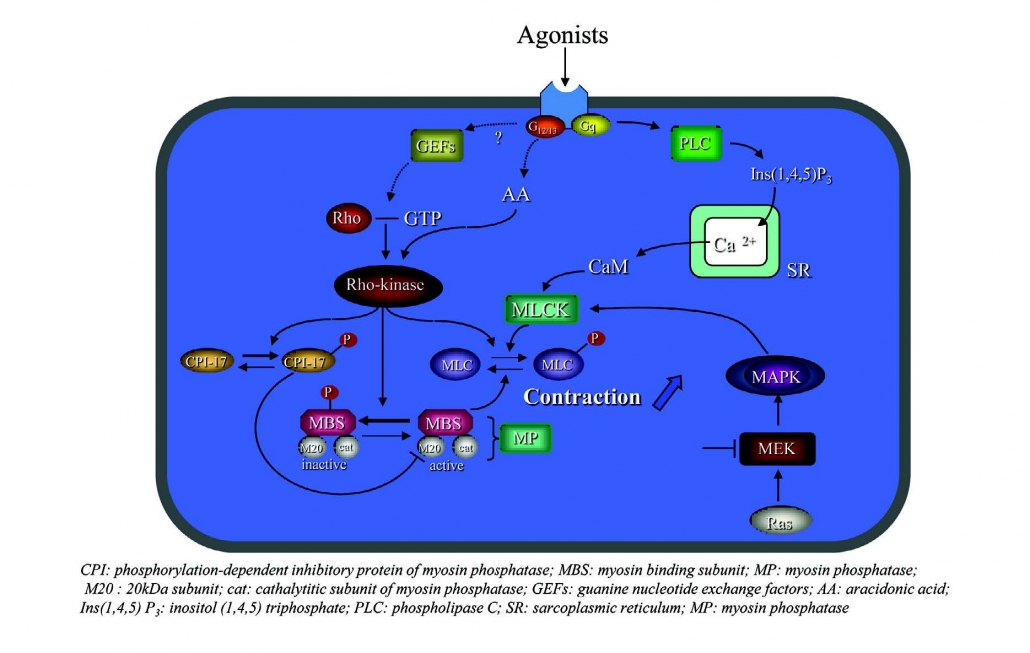
AitleIgnooui A, Hon W, Lamine F, Eutemene H, Fioramonti J, Bueno L, Theodorou V. Lactobacillus farciminis treatment suppresses stress induced visceral hypersensitivity: a possible action through interaction with epithelial cell cytoskeleton contraction. Farciminis乳杆菌治疗可抑制应激引起的内脏超敏反应:通过与上皮细胞细胞骨架收缩相互作用可能产生的作用Gut 2006; 55 :1090-4.
O’Mahony L, McCarthy J, Kelly P, Hurley G, Luo F, Chen K, O’Sullivan GC, Kiely B, Collins JK, Shamahan F, Quigley EM. Lactobacillus and bifidobacterium in irritable bowel syndrome: symptom responses and relationship to cytokine profiles, 肠易激综合征的乳杆菌和双歧杆菌:症状反应及其与细胞因子谱的关系 2005;128 :541-5.
Forsythe P, Wang B, Khambati I, Kunze WA. Systemic Effects of Ingested Lactobacillus Rhamnosus: Inhibition of Mast Cell Membrane Potassium (IKCa) Current and Degranulation. 摄入鼠李糖乳杆菌的全身作用:肥大细胞膜钾(IKCa)电流的抑制和脱粒PloS 2012;7(7): e41234.
Schiffer C, Lalanne AJ, Cassard L, Mancardi DA, Malbec O, Bruhns P, Dif F, Daeron M. A strain of Lactobacillus casei inhibits the effector phase of immune inflammation. 干酪乳杆菌菌株抑制免疫炎症的效应期 J Immunol. 2011; 187: 2646-55.
Blanco-miguez A, Gutierrez-jacome A, Fdez-riverola F, Lourenco A, Sanchez B. A peptidome-based phylogeny pipeline reveals differential peptides at the strain level within Bifidobacterium animalis subsp. Lactis. 基于肽组的系统发生管道揭示了动物双歧杆菌乳酸亚种内菌株水平的差异肽 Food Microbial. 2016; 60: 137-41.
Sorkar. Whether viable and dead probiotic ore equally efficacious?是否同样有效 Nutrition & Food Science. 2018.
Alander, M. Effect of galacto-oligosaccharide supplementation on human faecal microflora and on survival and persistence of Bifidobacterium lactis Bb-12 in the gastrointestinal tract. 补充低聚半乳糖对人粪便微生物区系以及乳酸双歧杆菌Bb-12在胃肠道中的存活和持久性的影响 Dairy J. 2001, 11, 817-825.
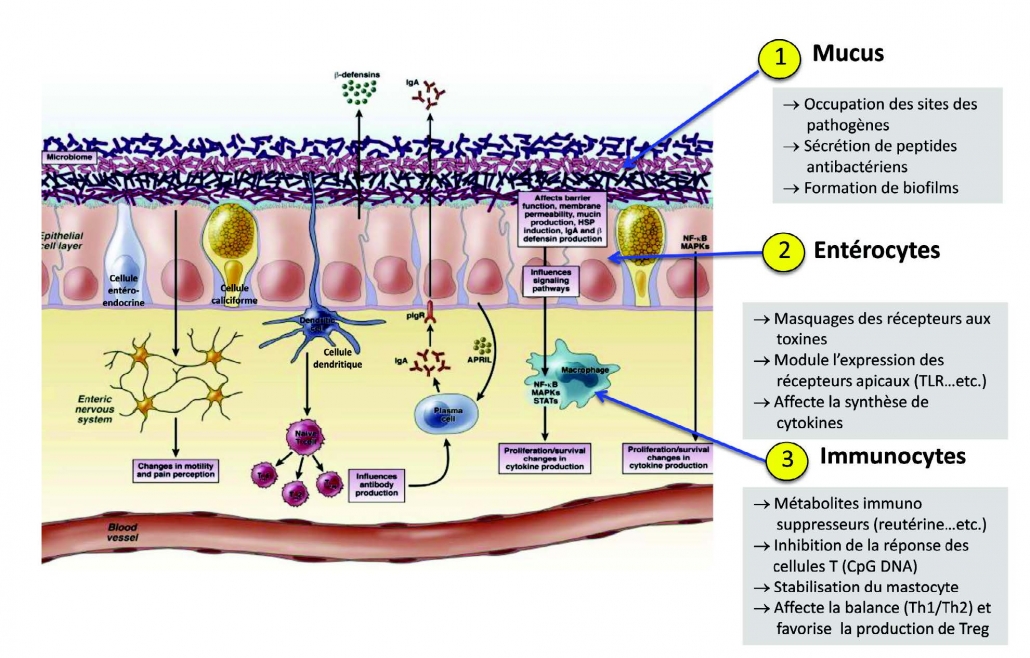
Ruiz I, Delgado S, Ruas Madiedo P, Sanchez B, Margolles A. Bifidobacteria and Their Molecular Communication with the Immune System. Front Mi 双歧杆菌及其与免疫系统的分子通讯2017 Dec 4;8:2345.
Oliveria LF, Salvador SL, Silva PH, Furlaneto FA, Figueiredo L, Casarin R, Ervolino E, Palioto DB, Souza SL, Taba M Jr, Navoes AB Jr, Messora MR. Benefits of Bifidobacterium animalis subsp. lactis Probiotic in Experimental Periodontitis动物双歧杆菌乳酸亚种益生菌在实验性牙周炎中的益处J Periodontology 2017 Feb;88(2):197-208.
Saavedra,M.; Bauman, N.A.; Oung, Perman, J.A; Yolken, R.H. Feeding of Bifidobacterium bifidum and Streptococcus thermophilus to infants in hospital for prevention of diarrhoea and shedding of rotavirus. 住院婴幼儿发育双歧杆菌和嗜热链球菌预防腹泻的的和轮状病毒脱落Lancet 1994, 344, 1046-1049.
Chouraqui, J.P.; van Egroo,; Fichot, M.C. Acidified milk formula supplemented with bifidobacterium lactis: impact on infant diarrhea in residential care settings补充乳酸双歧杆菌的酸化牛奶配方:对住院护理环境中婴儿腹泻的影响. J. Pediaitr. Gastroenterol. Nutr. 2004, 38, 288-292.
Lorenzo Morelli, Maria L Callegari, Vania Prebiotics, Probiotics, and Symbiotics: A Bifidobacterial View. The Bifidobacteria and Related Organisms. 益生元、益生菌和共生菌:双歧杆菌: 双歧杆菌和相关生物. Biology, Taxonomy, Applications. 2018, Pages 271-293.
Christian Riedel. Chapter 13 – Clinical Significance of Bifidobacteria. The Bifidobacteria and Related Organisms. 双歧杆菌的临床意义: 双歧杆菌及相关生物Biology, Taxonomy, Applications. 2018, Pages 221-234.
Kailasapathy K, Chin J. Survival and therapeutic potential of probiotic organisms with reference to Lactobacillus acidophilus and Bifidobacterium spp益生菌嗜酸乳杆菌和双歧杆菌属的存活和治疗潜力 Immunol Cel Biol 2000;78:80-88.
Gibson GR, Fuller R. Aspects of In Vitro and In Vivo Research Approaches Directed Toward Identifying Probiotics and Prebiotics for Human Use. 体外和体内研究结果:从各个方面定义供人类使用的益生菌和益生元J Nutr 2000; 130: 391s-395s.
Gupta P, Andrew H, Kirschner BS, Guanddini S. Is Lactobacillus GG helpful in children in Crohn’s disease? Results of a preliminary, open-label study. GG乳杆菌对克罗恩病患儿有帮助吗? 初步开放标签研究的结果。Pediatr Gastroenterol Nutr 2003; 31: 453-457.
Armuzzi Al, Cremonini F, Qjetti V, Bartolozzi F, Conduceci F, Candelli M, Santarelli L, Cammarota G, De Lorenzo A, Pola P, Gasbarrini G, Gasbarrini A. Effect of Lactobacillus GG supplementation on anfibio associated gastrointestinal side effects during Helicobacter Mori eradication therapy: a pilot study. 补充乳酸杆菌GG对森氏幽门螺杆菌根除治疗过程中与胃泌素相关的胃肠道副作用的影响:一项初步研究 2031; 63(1): 1-7.
Oberhelman RAI, Giknan RH, Sheen P, Taylor DN, Black RE, Cabrera L Lescano AG, Meza R, Madico G. A placebo-controled trial of lactobocillus GG to prevent diarrhea in undernourished Peruvian chil GG乳杆菌预防营养不良的秘鲁儿童腹泻的安慰剂对照试验1 Pediatr 1999;134(1): 15-20.
Nermes M, Kantele JM, Atosuo TJ, Salminen S, Isolauri E. Interaction of orally administered Lactobacillus rhamnosus GG with skin and gut microbiota and humoral immunity in infants with atopic dermatitis口服特应性鼠李糖乳杆菌GG与特应性皮炎婴儿的皮肤和肠道菌群及体液免疫的相互作用. Clin Exp Alergy. 2011 Mai ; 41(3) : 370-
Rautave S, Arvilommi H, Isolauri E. Specific probiotics in enhancing maturation of IgA responses in formula-fed infants特定的益生菌可提高配方奶粉婴儿IgA反应的成熟度. Pediatric Research (2006)60, 221-224.
Rautave S, Salminen S, Isolauri E. Specific probiotics in reducing the risk of acute infections in infancy–a randomised, double-blind, placebo-controlled study特定益生菌可降低婴儿急性感染的风险:一项随机双盲安慰剂对照的研究. British Journal of Nutrition (2009), 101,1722-26.
Morales A. Carvajal P, Siva N al. Clinical Effects of Lactobacillus rhamnosus in Non-Surgical Treatment of Chronic Periodontitis: A Randomized Placebo-Controlled Trial With 1-Year Follow-Up鼠李糖乳杆菌在非手术治疗慢性牙周炎中的临床效果:一项为期1年随访的随机安慰剂对照试验. J Periodontal 2016; 4:1-12.
Pier Sandro Cocconcell Istituto di microbiologia-Centro Ricerche biotecnologiche. Unnersita Cattolica del sacro cuore 2007.
E. Sanders, Invited review: the scientific basis of Lactobacillus acidophilus NCFM functionality as a probiotic特邀评论:嗜酸乳杆菌NCFM作为益生菌的功能的科学基础. J. Dairy Sci. 84; 201: 319-331.
Wildt SI, Munck LK, Vinter-Jensen L Hanse BF, Nordgaard-Lassen I, Christensen S, Avnstroem S, Rasmussen SN, Rumessen JJ. Probiotic treatment of collagenous colitis: a randomized, double-blind, placebo-controlled trial with Lactobacillus acidophilus and Bifidobacterium animalis subsp. Lactis益生菌治疗胶原性结肠炎:嗜酸乳杆菌和动物双歧杆菌乳亚种的随机双盲安慰剂对照试验. Inflamm Bowel Dis. 2006;12 (5):395-401.
Chatteriee S, Kar P, Das T, Ray S, Gangulyt S, Raiendiron C, Mitra M,. Randomised placebo-controlled double blind multicentric trial on efficacy and safety of Lactobacillus acidophilus LA-5 and Bifidobacterium BB-12 for prevention of antibiotic-associated diarrhoea嗜酸乳杆菌LA-5和双歧杆菌BB-12预防与抗生素相关的腹泻的功效和安全性的随机安慰剂对照双盲多中心试验. J Assoc Physicians India 2013. oct; 61(10): 708-12.
Black, FT., Laulund, S. A study on the recovery of ingested, encapsulated Lactobacillus acidophilus and Bifidobacterium bifidum from duodenal fluid and faeces. 从十二指肠液和粪便中回收包封的嗜酸乳杆菌和双歧双歧杆菌的研究 Chr. Hansen Internal Report.
Black, FT., Einarsson, K., Lidbeck, A, Orrhage, K., Nord, C.E. Effect of lactic acid producing bacteria on the human intestinal microflora during ampicillin treatment. 产乳酸菌对氨苄西林治疗期间人肠道菌群的影响 Scand. J. Infect. Dis., 23: 247-254
Maaike C. de Vries. Lactobacillus plantarum-survival, functional and potential probiotic properties in the human intestinal tract. 植物乳杆菌在人肠道中的存活、功能和潜在益生菌性质International Dairy Journal 2006 ; 16: 1018-1028.
Vilahur GI, Lopez-Bernal S, Camino S, Mendiata G, Padro T, Badimon L. Lactobacillus plantarum CECT 7315/7316 intake modulates the acute and chronic innate inflammatory response植物乳杆菌CECT 7315/7316的摄入量可调节急性和慢性先天性炎症反应. Eur J Nutr. 2014 Nov
Li Cl, Nie SP, Zhu KX, Ding Q, Li C, Xiong T, Xie Lactobacillus plantarum NCU116 improves liver function, oxidative stress and lipid metabolism in rats with high fat diet induced non-alcoholic fatty liver disease. 植物乳杆菌NCU116改善高脂饮食诱发的非酒精性脂肪肝大鼠的肝功能、氧化应激和脂质代谢Food Funct 2014 Nov 19;5: 3216-23.
Wang LX, Liu K, Gao DW, Hoo JK. Protective effects of two Lactobacillus plantarum strains in hyperlipidemic mice. 两种植物乳杆菌对高脂血症小鼠的保护作用World J Gastroenterol. 2013:19 :3150-6.
Wharwell PJ, Altringer L, Morel J, Bond Y, Charbonneau D, O‘mahony L, Kiely B, Shanahan F, Quigley EM. Efficacy of an encapsulated probiotic Bifidobacterium infantis 35624 in women with irritable bowel syndrome封装的益生菌婴儿双歧杆菌35624在女性肠易激综合征中的功效. Am J Gastroenterol. 2006 l; 101(7): 1581-90.
McKernan DP, Fizgerald P, Dinan TG, Cryan JF. The probiotic Bifidobacterium infantis 35624 displays visceral antinociceptive effects in the rat. 益生菌婴儿双歧杆菌35624个显示内脏大鼠镇痛作用Neurogastroenterol Motil. 2010; 22(9):1029-
O’Mahony L, McCarthy J, Kelly P et Lactobacillus and bifidobacterium in irritable bowel syndrome: symptom responses and relationship to cytokine profiles. 肠易激综合症中的乳酸杆菌和双歧杆菌:症状反应及其与细胞因子谱的关系Gostroenterology 2005; 128: 541-51.
Vivatvakin B, Kowitdamrong E. Randomized control trial of live Lactobacillus acidophilus plus Bifidobacterium infantis in treatment of infantile acute watery diarrhea嗜酸乳杆菌联合婴儿双歧杆菌治疗婴儿急性水样腹泻的随机对照试. J Med Assoc Thai. 2006 Sep ; 89 Suppl 3 : S126-33.
Gabriela Sinkiewi Lactobacillus reuteri in heahh and desease罗伊氏乳杆菌在健康和疾病中的作用, Doctoral dissertations, Malmo university, 2010.
Lactobacillus reuteri as a therapeutic agent in acute diarrhea in young children 罗伊氏乳杆菌作为幼儿急性腹泻的治疗剂. Journal of pediatric gastroenterology and nutrition. J Pediair Gastroenterol Nutr. 1997 Apr; 24(4): 399-404.
Savino , Pelle E., Palumeri E., Oggero R. and Miniero R. (2007). ‘ Lactobacillus reuteri (American Type Culture Collection Strain 55730) versus simethicone in the treatment of infantile colic: a prospective randomized study罗伊氏乳杆菌(美国典型培养物保藏菌株55730)与西甲硅油治疗婴儿绞痛的前瞻性随机研究. Pediatrics 119 (1): 124-130.
Cosas LA, Edens FW, Dobragasz WJ. Lactobacillus reuteri: an effective probiotic for poultry and other animals. 罗伊氏乳杆菌:对家禽和其他动物有效的益生菌。Lactic acid bacteria, 2nd ed. New York Marcel Dekker, 1998: 475-518.
Alak JL, Wolf BW, Mdurvwa EG, Pimentel-Smith GE, Adeyemo O. Effect of Lactobacillus reuteri on intestinal resistance to Cryptosporidium parvum infection in a murine model of acquired immunodeficiency syndrome罗伊氏乳杆菌对获得性免疫缺陷综合症小鼠模型肠道抵抗小隐孢子虫感染的影响. J Infect Dis 1997; 175: 218-21.
Gruner D, Pais S and Schwendick F. 2016. Probiotics for managing caries and periodontitis: Systematic review and meta-analysis益生菌治疗龋齿和牙周炎:系统评价和荟萃分析. Journal of Dentistry. 48: 16-
Szkaradkiewicz AK, Stopa J, Karpinski TM. 2014. Effect of oral administration involving a probiotic strain of Lactobacillus reuteri on pro-inflammatory cytokine response in patients with chronic periodontitis口服罗伊氏乳杆菌益生菌菌株对慢性牙周炎患者促炎细胞因子反应的影响. Arch Immunol Ther Exp (Warsz). 62(6): 495-500.
Zang J, Li YQ, Zuo XL, Zhen YB, Yang J, Liu CH. Clinical trial: effect of active lactic acid bacteria on mucosal barrier function in patients with diarrhoea‐predominant irritable bowel syndrome. 临床试验:活性乳酸菌对腹泻型肠易激综合征患者黏膜屏障功能的影响 Aliment Pharmacol Ther. 2008;28:994-1002.
Jiang T, Mustapha A, Savaiano DA. Improvement of lactose digestion in humans by ingestion of unfermented milk containing Bifidobacterium longum. 摄入含有长双歧杆菌的未发酵乳可以改善人的乳糖消化J Dairy Sci. 1996; 79 :750-7.
Al-Sheraji SH, Ismail A, Manap MY, Mustafa S, Yusof RM, Hassan FA. Hypocholesterolaemic effect of yoghurt containing Bifidobacterium pseudocatenulatum G4 or Bifidobacterium longum BB536. 含假双歧杆菌G4或长双歧杆菌BB536的酸奶的降胆固醇作用Food Chem. 2012; 135: 356-
Rodriguez C, Medici M, Mozzi F, Fort de Valdez G. Therapeutic effect of Streptococcus thermophilus CRL 1190-fermented milk on chronic gastritis. 嗜热链球菌CRL 1190发酵乳对慢性胃炎的治疗作用World J Gastroenterol. 2010;16(13): 1622-
Shimosato T, Tohno M, Sato T, Nishimura J, Kawai Y, Saito T, Kitazawa H. Identification of a potent immunostimulatory oligodeoxynucleotide from Streptococcus thermophilus lacZ. 从嗜热链球菌lacZ鉴定有效的免疫刺激性寡脱氧核苷酸Anim Sci J. 2009; 80: 597-604.
Jain PK, McNaught CE, Anderson AD, MacFie J, Mitchell CJ. Influence of synbiotic containing Lactobacillus acidophilus La5, Bifidobacterium lactis Bb 12, Streptococcus thermophilus, Lactobacillus bulgaricus and oligofructose on gut barrier function and sepsis in critically ill patients: a randomised controlled trial. 含有嗜酸乳杆菌La5,乳酸双歧杆菌Bb 12,嗜热链球菌,保加利亚乳杆菌和低聚果糖的合生素对危重患者肠道屏障功能和败血症的影响:一项随机对照试验Clin Nutr. 2004 Aug; 23(4): 467-75.
Hickson M, D’Souza AL, Muthu N, Rogers TR, Want S, Rajkumar C, Bulpitt C Use of probiotic Lactobacillus preparation to prevent diarrhoea associated with antibiotics: randomised double blind placebo controlled trial. 益生菌乳酸菌制剂用于预防与抗生素有关的腹泻:随机双盲安慰剂对照试验BMJ. 2007 Jul 14;335(7610): 80.
Gionchetti P, Rizzello F, Morselli C, Poggioli G. Tombasco R, Colabrese C, Brigidi P, Vitali B, Straforini G, Campieri M. High-dose probiotics for the treatment of active pouchitis. 高剂量的益生菌治疗活性囊炎Dis Colon Rectum. 2007 Dec; 50(12): 2075-82; discussion 2082-2084.
Aso Y, Akaza H, Kotake T, Tsukamoto T, Imai K, Naito S. Preventive effect of a Lactobacillus casei preparation on the recurrence of superficial bladder cancer in a double-blind trial. The BLP Study Group. 在一项双盲试验中,干酪乳杆菌制剂对浅表性膀胱癌复发的预防作用。 BLP研究组Eur Urol. 1995; 27(2): 104-109.
Sachdeva A, Nagpal J. Effect of fermented milk-based probiotic preparations on Helicobacter pylori eradication: a systematic review and meta-analysis of randomized-controlled trials. 发酵乳基益生菌制剂对幽门螺杆菌根除的影响:随机对照试验的系统评价和荟萃分析European Journal of Gastroenterology & Hepatology. 2009; 21: 45-53.
Matar C, Theriault C, Perdigon G. Effects of milk fermented by Lactobacillus helveticus R389 on immune cells associated to mammary glands in normal and a breast cancer model. 瑞士乳杆菌R389发酵的牛奶对正常和乳腺癌模型中与乳腺相关的免疫细胞的影响 Immunobiology 2005;210(5):349-58
Griffiths M. V., Tellez A. M. Lactobacillus helveticus: the proteolytic system. 瑞士乳杆菌:蛋白水解系统 Microbiol. 4:30 10.3389/fmicb.2013.
Taverniti, Guglielmetti S. Heahl-promoting properties of Lactobacillus helveticus. 瑞士乳杆菌的健康促进特性Front. Microbial. 3:392 10.3399/frnicb.2012.
K Hatakka, AJ Ahola, H Yli-Knuuttila. Probiotics reduce the prevalence of oral candida in the elderly–a randomized controlled trial. 益生菌可降低老年人口服念珠菌的患病率(一项随机对照试验)Journal of Dental Research, 2007, 86(2):125-130
Ishikawa H, et al. Suppression of Periodontal Pathogenic Bacteria in the Saliva of Humans by the Administration of Lactobacillus salivarius TI2711. 唾液乳杆菌TI2711对人唾液中牙周病原细菌的抑制作用Journal of the Japanese Society of Periodontology. 2003. 45(1):105-112
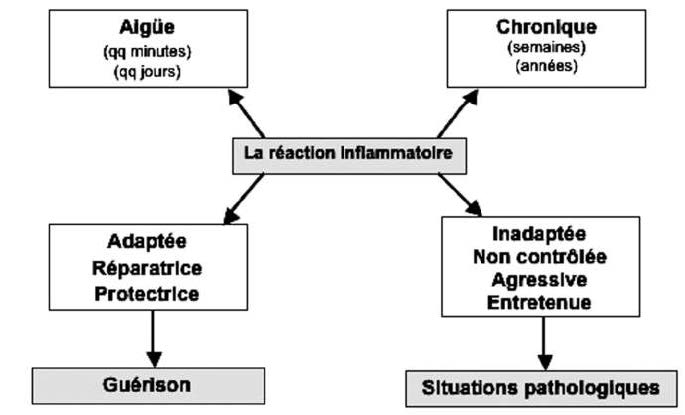
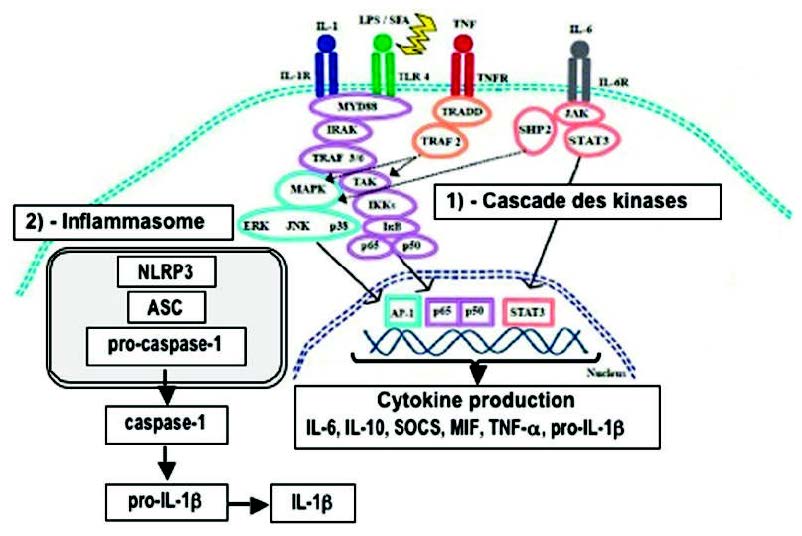
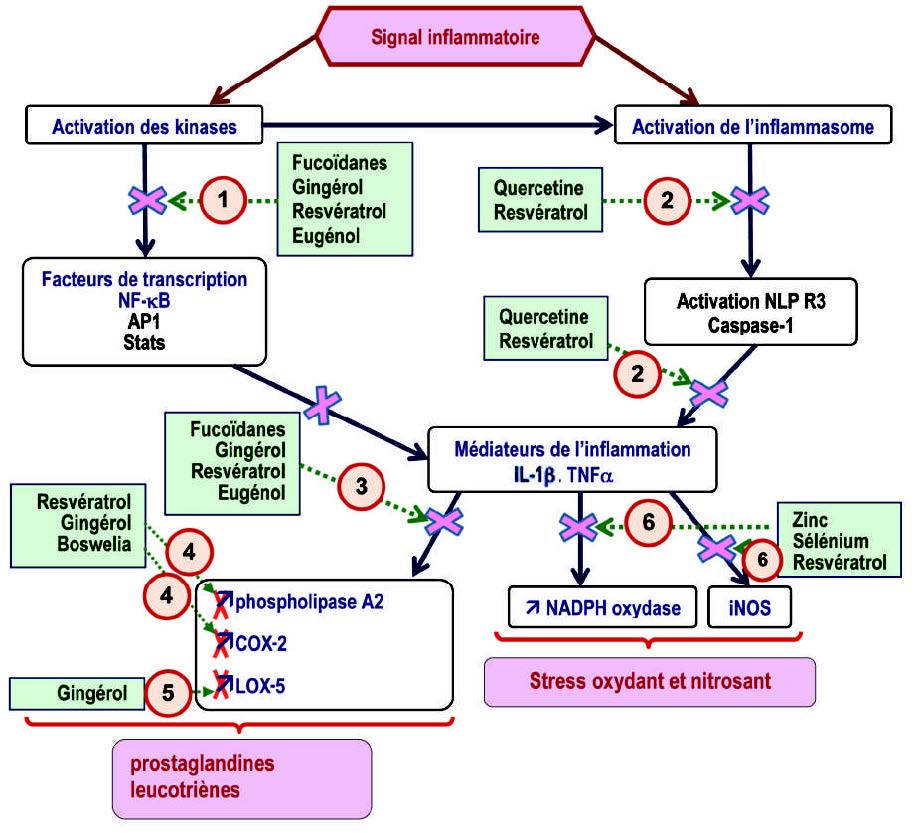
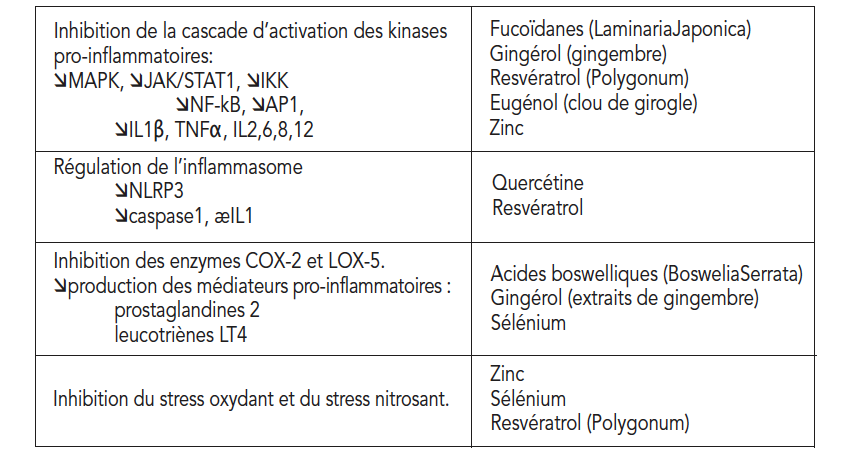
炎性小体是由几种蛋白质组成的膜复合物,包括胱天蛋白酶1,胱天蛋白酶1在识别炎症信号后被蛋白质家族NLRP(Nod Like ReceptorProtein)激活。 迄今为止,已经描述了各种炎症小体,包括临床上与肥胖、胰岛素抵抗和轻度炎症有关的NLRP3(7)。 NLRP3半胱天冬酶1负责从无活性的前IL1β形成IL1β。抑制NLRP3是抗轻度炎症的优先选项。
Moore N et al. Adverse drug reactions and drug-drug interactions with over-the-counter NAIDs. 与非处方非甾体抗炎药的不良药物反应和药物相互作用 Ther Clin Risk Manag. 2015; 11:1061-75.
Patterson H et al. Protein kinase inhibitors in the treatment of inflammatory and autoimmune diseases. 蛋白激酶抑制剂在炎性和自身免疫性疾病的治疗中的使用Clin Exp Immunol. 2013; 176:1-10.
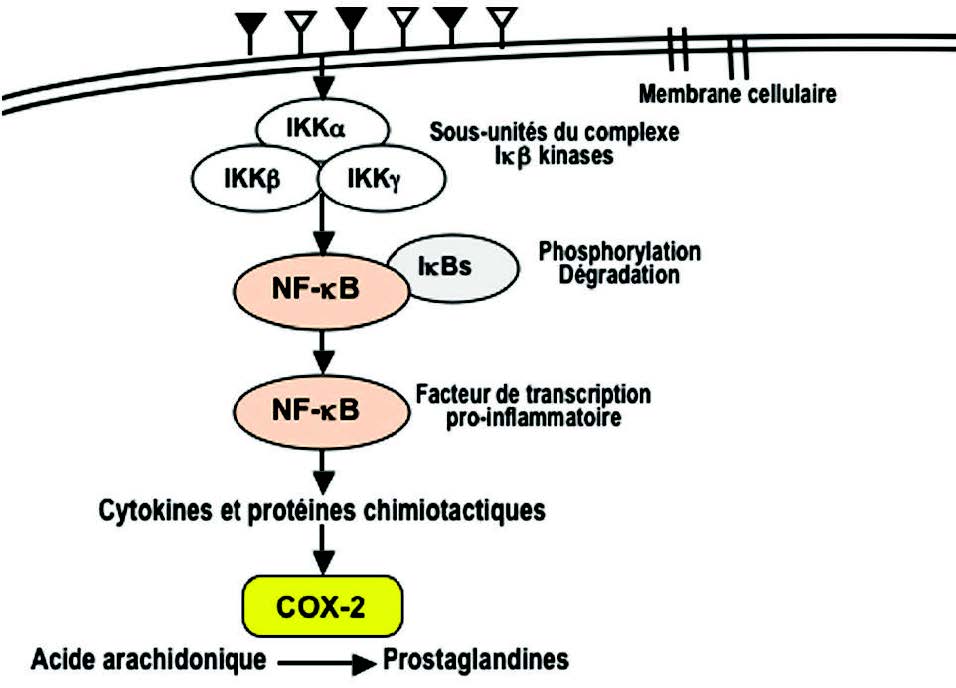
Hinz M et al. the IkBkinase complex in NF-kB regulation and beyond; IkB激酶复合物在NF-kB调控中的作用 EMBO rep. 2014; 15(1):46-61.
Ledoux AC et al. NF-kB and the cell cycle. NF-kB与细胞周期Biochem Soc Trans. 2014; 42(1):76-81.
Latz E et al. Activation and regulation of the inflammasomes. 炎症小体的活化和调节 Nat Rev Immunol; 2013; 13(6).
Lamkanfi M et al. Mechanisms and functions of inflammasomes. 炎症小体的机制和功能Cell. 2014; 157:1013-1022.
Abderrazak A et al. NLRP3 inflammasome: From a danger signal sensor to a regulatory node of oxidative stress and inflammatory diseases. NLRP3炎症小体:从危险信号传感器到氧化应激和炎症性疾病的调节节点Redox Biol. 2015; 4:296-307.
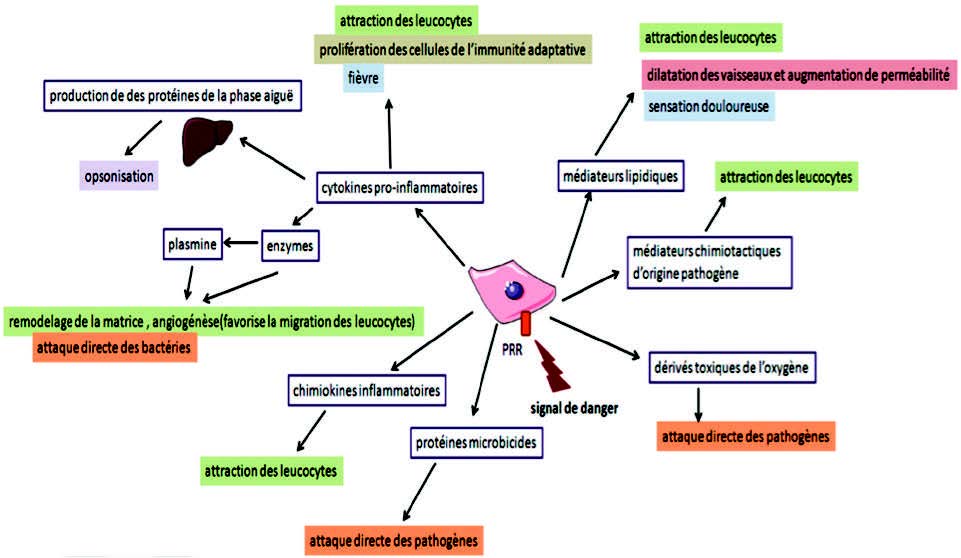
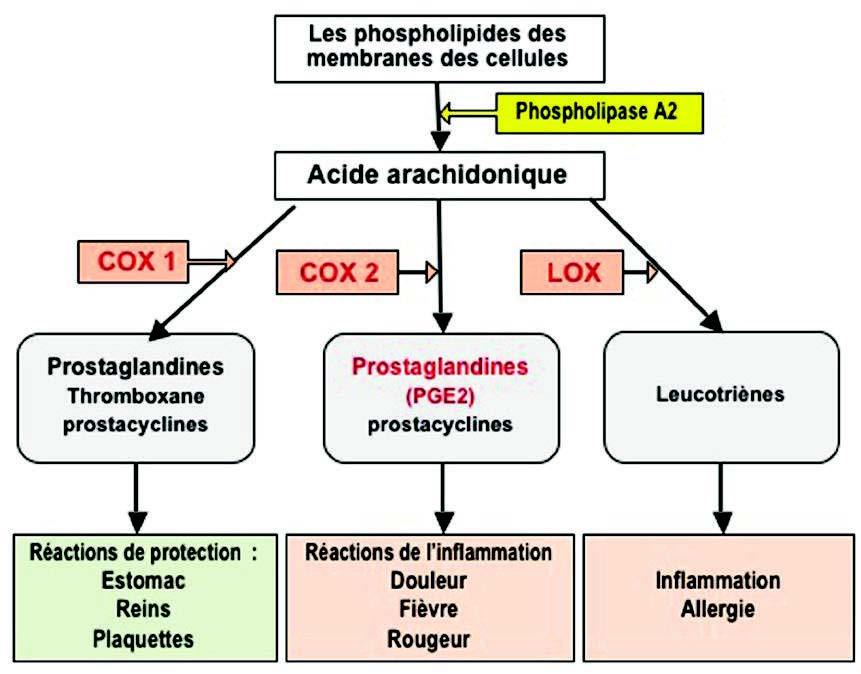
Henrotin Y et al. Les médiateurs biochimiques de l’inflammation. 炎症的生化介质Rev Med Liege. 2001; 56:6:433-442.
Lawrence T. The nuclear Factor NF-kB pathway in inflammation. 炎症中的核因子NF-kB途径 Cold Spring Harb Perspect Biol. 2009; 1(6).
Minihane AM et al. Low-grade inflammation, diet composition and health: current research evidence and its translation. 轻度炎症、饮食组成和健康:最新研究证据及其翻译Br J Nutr. 2015; Jul 31:1-14.
Barbara G et al. A role for inflammation in irritable bowel syndrome? 炎症在肠易激综合症中起作用?Gut 2002; 51 Suppl 1:i41-4.
Chen L et al. Mechanisms linking inflammation to insulin resistance. 炎症与胰岛素抵抗相关的机制 Int J Endocrinol. 2015; 2015:508409.
Zalli A et al. Low-grade inflammation predicts persistence of depressive symptoms. 轻度炎症预示着抑郁症状的持续存在 Psychopharmacology. 2015 ; Apr16.
Singh A, karrar S. The role of intracellular organisms in the pathogenesis of inflammatory arthritis. 细胞内生物在炎症性关节炎发病中的作用 Int J Inflam. 2014; 2014:158793.
Podzimek S et al. C reactive protein in peripheral blood of patients with chronic and aggressive periodontitis, gingivitis and gingival recessions; 性和侵袭性牙周炎、牙龈炎和牙龈退缩的患者外周血C反应蛋白Mediators Inflamm. 2015; 2015:564858 .
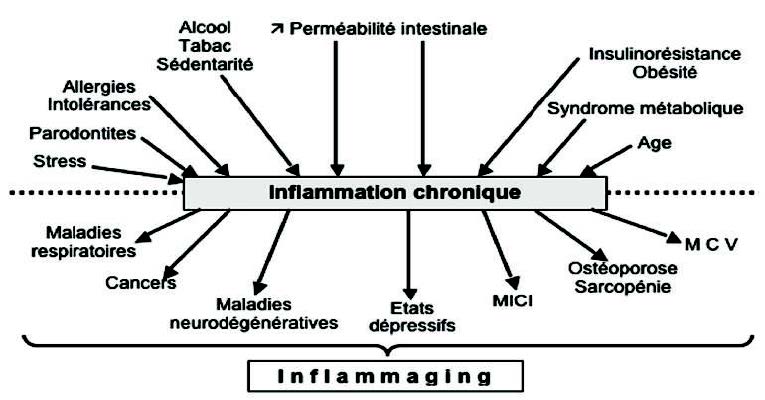
Baylis D et AL understanding how we age: Insights into inflammaging. 洞悉炎症 Longev Health Span. 2013; 2(1):8.
Thornburn AN et al. Diet, metabolites and western life style inflammatory diseases. 饮食、代谢物和西方生活方式的炎症性疾病Immunity. 2014; 40(6):833-42.
Lopez-Alarcon M et al. Excessive refined carbohydrates and scarce micronutrients intakes increase inflammatory mediators and insulin resistance in prepubertal and pubertal obeses children independently of obesity. 青春期前和青春期肥胖儿童中过量摄入的精制碳水化合物和微量营养素摄入不足会增加炎症介质和胰岛素抵抗,而与肥胖无关Mediators Inflamm. 2014; 2014:849031.
Aldaham S et al. Smoking status effect on inflammatory markers in a randomized trial of current and former heavy smokers. 在现有和以前的重度吸烟者的随机试验中,吸烟状态对炎症标志物的影响 Int J Inflam. 2015; 2015:439396.
Troester N et al. Sleep related breathing disorders and inflammation. The missing link? A cohort study evaluating the interaction of inflammation and sleep related breathing disorders and effects of treatments. 睡眠相关的呼吸障碍和炎症。缺少相关性? 一项队列研究,评估炎症与睡眠相关的呼吸障碍的相互作用以及治疗效果 PLoS One. 2015; 10(9):e0137594.
Pruimboom L et al. Physical activity protects against metabolic stress induced by a post prandial and chronic inflammation. 体育锻炼可防止餐后和慢性炎症引起的代谢压力. Behav Neurol. 2015; 2015:569869
Gouin JP et al. Chronic stress, daily stressors, and circulating inflammatory markers. 慢性压力、日常压力和循环炎症标记 Health Psychol. 2012; 31(2):264-8.
Mostafavi et al. Inflammatory markers in relation to long term air pollution. 与长期空气污染有关的炎症标志 Environ. Int. 2015; 81:1-7.
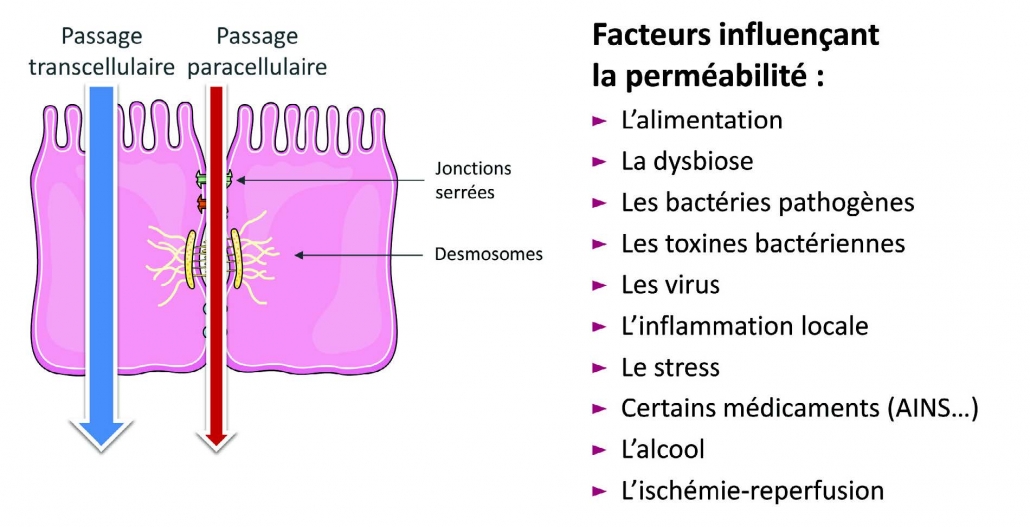
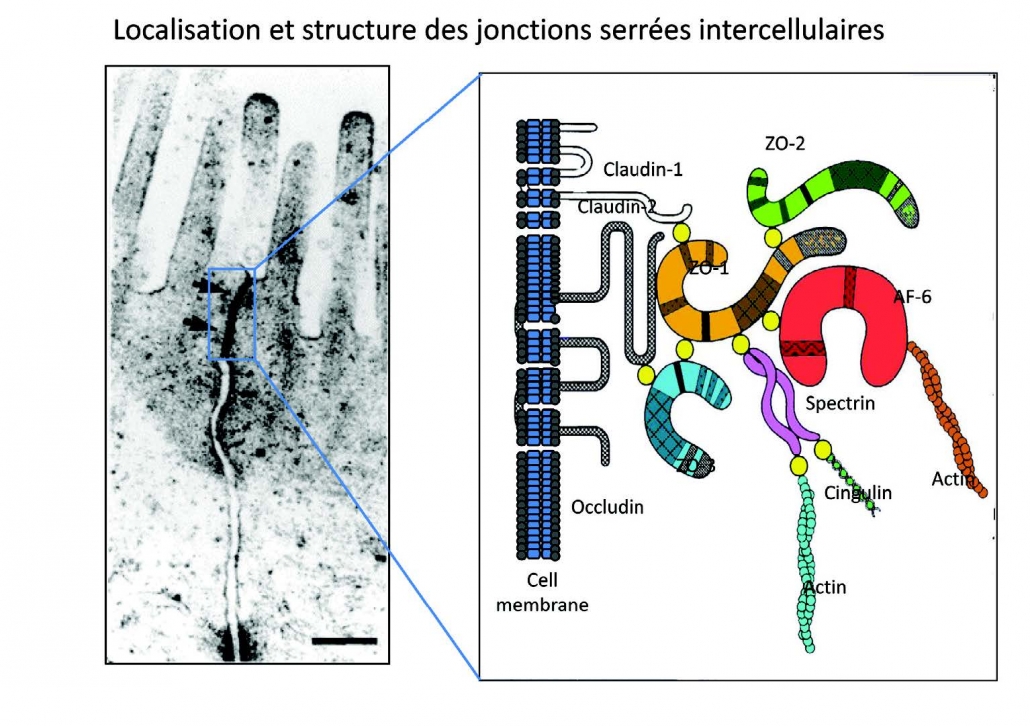
Sung HL; Intestinal permeability regulation by tight junction: implication on inflammatory bowel diseases. 紧密连接调节肠道通透性:对炎症性肠病的影响 Intest Res. 2015; 13(1):11-18.
Shaw AC et al. Age-dependent dysregulation of innate immunity. 先天性免疫的年龄依赖性失调 Nature Rev Immunol. 2013; 13,875-887.
Franceschi C, Campisi J. Chronic inflammation (inflammaging) and its potential contribution to ag-associated diseases. 慢性炎症(发炎)及其与年龄相关疾病的潜在贡献 J Gerontol A Biol Sci Med Sci. 2014; 69 Suppl 1: S4-9.
Apostopoulos V et al. The complex immunological and inflammatory network of adipose tissue in obesity.肥胖中脂肪组织的复杂免疫和炎症网络 Mol Nutr Food Res. 2015 September.
Izaola O et al. Inflammation and obesity (lipoinflammation). 炎症和肥胖(脂肪炎症)Nutr Hosp. 2015; 31(6):2352-8.
Dalmas E et al. T cell-derived IL22 amplifies IL1beta driven inflammation in human adipose tissue: relevance to obesity and type 2 diabetes. T细胞衍生的IL22放大人脂肪组织中IL1beta驱动的炎症:与肥胖和2型糖尿病的相关性 Diabetes. 2014; 63(6):1966-77.
Neels JG. A role for 5-lipoxygenase products in obesity-associated inflammation and insulin resistance. 5-脂氧合酶产品在肥胖相关的炎症和胰岛素抵抗中的作用 Adipocyte. 2013; 2(4):262-5.
Leon-Cabrera S et al. Hyperleptinemia is associated with parameters of low-grade systemic inflammation and metabolic dysfunction in obese human beings. 高瘦素血症与肥胖者的轻度全身性炎症和代谢功能障碍相关 Front Integr Neurosci. 2013; 7:62.
Munhoz CD et al. Stress-induced neuro-inflammation: mechanisms and new pharmacological targets. 应激诱导的神经炎症:机制和新的药理目标 Braz J Med Biol Res. 2008; 41(2):1037-46.
Berk M et al. So depression is an anti-inflammatory disease, but when does the inflammation come from? 抑郁症是一种抗炎性疾病,但是炎症是什么时候产生的呢?BMC Med; 2013; 12:200.
Kontzias A et al. Kinase inhibitors in the treatment of immune-mediated disease 激酶抑制剂在免疫介导疾病的治疗中; Med Rep. 2012; 4:5.
Asadullah K et al. Protein kinase inhibitors for the treatment of inflammation- an overview. 蛋白激酶抑制剂治疗炎症-概述 Antiinflamm Antiallergy Agents Med Chem. 2007; 6:3-4.
Fitton JH. Therapies from fucoidan: multifunctional marine polymers 岩藻依聚糖的治疗方法:多功能海洋聚合物; Mar Drugs. 2011; 9(10):1731-1760.
Cumashi A et al. A comparative study of the anti-inflammatory, and antiadhesive activities of nine different fucoidans from brown seaweeds. 九种褐藻海藻岩藻聚糖的抗炎和抗黏附活性的比较研究 Glycobiology. 2007; 17:541-552.
Park HY et al. Antiinflammatory effects of fucoidan through inhibition of NF-kB, MAPK and AKT activation in LPS-induced BV2 microglia cells. 岩藻依聚糖通过抑制LPS诱导的BV2小胶质细胞的NF-kB,MAPK和AKT活化而具有抗炎作用 Food Chem Toxicol. 2011; 49(8):1745-52.
Kim KJ et al. Fucoidan, a sulfated polysaccharide, inhibits adipogenesis through the mitogen-activated protein kinase pathway in 3T-L1 preadipocytes. 岩藻依聚糖是一种硫酸化多糖,可通过3T-L1前脂肪细胞中的促分裂原活化蛋白激酶途径抑制脂肪形成。Life Sci; 2010; 86(21-22):791-7.
Religa P et al. Fucoidan inhibits smooth muscle cell proliferation and reduces MAPK activity. 岩藻依聚糖抑制平滑肌细胞增殖并降低MAPK活性 Eur J Vasc Endovasc Surg. 2000; 20(5):419-26.
Van Breemen RB et al. Cyclooxygenase-2 inhibitors in ginger (Zingiber officinale). 生姜中的环氧合酶2抑制剂(姜)Fitoterapia. 2011; 82(1):38-43.
Grzanna R et al. Ginger, an herbal medicinal product with broad anti-inflammatory actions. 生姜,具有广泛的抗炎作用的草本植物 J Med Food 2005; 8(2):125-132.
Lee TY et al. 6 gingerol inhibits ROS and iNOS through the suppression of PKC-alpha and NFkappaB pathways in LPS stimulated mouse macrophage. 6-姜油醇通过抑制LPS刺激的小鼠巨噬细胞中的PKC-alpha和NFkappaB途径抑制ROS和iNOS Biochem Biophys Res Comm; 2009; 382(1):134-139.
Wang S et al. Biological properties of 6-gingerol: a brief review 6-姜油醇的生物学特性:简要评述. Nat Prod Commun. 2014; 9(7):1027-30.
Singletary K. Ginger, an overview of health benefits. 生姜,对健康的益处概述 Nutrition Today. 2010; 45:4,171-183.
Al-Nahain A et al. Zingiber officinale: A potential plant against rheumatoid arthritis; 生姜:一种有抗风湿性关节炎潜力的植物Arthritis 2014; 2014:159089.
Atashak S et al. Obesity –related cardiovascular risk factors after long-term resistance training and ginger supplementation; 肥胖 -长期阻力训练和补充生姜相关的心血管危险因素 J sports Sci Med. 2011; 10(4):685-91.
Mahluji S et al. Antiinflammatory effects of Zingiber officinale in type 2 diabetic patients. 生姜对2型糖尿病患者的抗炎作用 Adv Pharm Bull. 2013; 3(2):273-6.
Arablou T et al. The effect of ginger consumption on glycemic status, lipid profile, and some inflammatory markers in patients with type 2 diabetes mellitus. 摄入生姜对2型糖尿病患者血糖状态、血脂和某些炎症指标的影响 Int J Food Sci. 2014; 65(4):515-20.
Wu J et al. Quercetin, luteolin, alleviate TXNIP and NLRP3-mediated inflammation and apoptosis with regulation of AMPK in endothelial cells. 槲皮素、木犀草素通过调节内皮细胞中的AMPK减轻TXNIP和NLRP3介导的炎症和细胞凋亡 Eur J Pharmacol. 2014; 745:59-68.
Vasquez Prieto MA et al. Catechin and quercetin attenuate adipose inflammation in 3T3-L1 adipocytes. 儿茶素和槲皮素可减轻3T3-L1脂肪细胞中的脂肪炎症 Mol Nutr Food Res. 2015; 59(4):622-33.
Dower JI et al. Supplementation of the pure flavonoïds epicatechin and quercetin affects some biomarkers of endothelial dysfunction and inflammation in hypertensive adults: a randomized double-blind placebo controlled crossover trial补充纯黄酮表儿茶素和槲皮素会影响高血压成年人内皮功能障碍和炎症的某些生物标志物:一项随机双盲安慰剂对照交叉试验. J Nutr 2015; 145(7):1459-63.
Svajger U et Jeras M. Antiinflammatory effects of resveratrol and its potential use in therapy of immune-mediated diseases. 白藜芦醇的抗炎作用及其在免疫介导疾病治疗中的潜在应用 Int Rev Immunol. 2012; 31(3):202-22.
Bollmann F et al. Resveratrol post-transcriptionally regulates pro-inflammatory gene expression via regulation of KSRP RNA binding activity. 白藜芦醇在转录后通过调节KSRP RNA结合活性来调节促炎基因的表达 Nucleic acids Res. 2014; 40(20):12555-69.
Chang YP et al. Resveratrol inhibits NLRP3 inflammasome activation by preserving mitochondrial integrity and augmenting autophagy. 白藜芦醇通过保持线粒体完整性并增强自噬来抑制NLRP3炎性体激活 J Cell Physiol. 2015; 230(7):1567-79.
Ren Z et al. Resveratrol inhibits NFkB signaling through suppression of p65 and IkB kinase activities. Pharmazie. 2013; 68(8):689-94.
Ghanim H et al. An anti-inflammatory and reactive oxygen species suppressive effects of an extract of polygonum cuspidatum containing resveratrol. 白藜芦醇通过抑制p65和IkB激酶活性抑制NFkB信号传导 J Clin Endocrinol Metab. 2010; 95(9):E 1-8.
Tome-Carneiro J et al. One-year supplementation with a grape extract containing resveratrol modulates inflammatory-related microRNAs and cytokines expression in peripheral blood mononuclear cells of type 2 diabetes and hypertensive patients with coronary artery disease. 一年补充含有白藜芦醇的葡萄提取物可调节2型糖尿病和冠心病高血压患者外周血单个核细胞中炎症相关的microRNA和细胞因子的表达 Pharmacol Res. 2013; 72:69-82.
Ghanim H et al. A resveratrol preparation suppresses oxidative and inflammatory stress response to a high fat high carbohydrate meal. 白藜芦醇制剂可抑制对高脂高碳水化合物膳食的氧化和炎症应激反应 J Clin Endocrinol Metab. 2011; 96(5):1409-14.
Yeh JL et al. Eugenol suppress LPS induced iNOS expression by down regulating NF-kappa B and AP1 through inhibition of MAPKs and AKT/IkBα signaling pathways in macrophages. 丁香油酚通过抑制巨噬细胞中的MAPKs和AKT /IkBα信号通路来下调NF-κB和AP1,从而抑制LPS诱导的iNOS表达 Int J Immunopathol Pharmacol. 2011; 24(2):345-56.
Pan C et al. Antiasthmatic effects of eugenol in a mouse model of allergic asthma by regulation of NF-kB pathway; 丁香油酚通过调节NF-kB途径对过敏性哮喘小鼠的哮喘作用 inflammation. 2015; 38(4):1385-93.
Abd El Motteleb DM et al. Differential effects of eugenol against hepatic inflammation and overall damage. 丁香酚对肝脏炎症和整体损伤的差异作用 J immunotoxicol. 2014; 11(3):238-45.
Kaur G et Sultana S. Evaluation of antiarthritic activity of isoeugenol in adjuvant induced arthritis in murine model. 异丁香酚在佐剂性关节炎小鼠模型中的抗关节炎活性评估 Food Chem Toxicol. 2012; 50(8):2869-95.
Siddiqui MZ. Boswellia serrata, a potential anti-inflammatory agent: an overview. 概述:齿叶乳香作为潜在的抗炎药 Indian J Pharm Sci. 2011; 73(3):255-61.
Catanzaro D et al. Boswellia serrata preserves intestinal epithelial barrier from oxidative and inflammatory damage. 齿叶乳香可保护肠上皮屏障免受氧化和炎性损害 PLoSOne 2015; 10(5):e0125375.
Sengupta K el al. A double blind, randomized placebo controlled study of the efficacy and safety of 5-loxin for treatment of osteoarthritis of the knee. 5-loxin治疗膝盖骨关节炎的有效性和安全性的双盲随机安慰剂对照研究。Arthritis Res Ther. 2008; 10(4):R85.
Gupta I et al. Effects of gum resin of Boswellia serrata in patients with chronic colitis. 齿叶乳香树脂在慢性结肠炎患者中的作用Planta Med. 2001; 67(5):391-5.
Gupta I et al. Effects of gum resin of Boswellia serrata in patients with bronchial asthma: results of a double blind, placebo controlled, 6 weeks clinically study. 齿叶乳香树脂对支气管哮喘患者的作用:双盲,安慰剂对照,6周临床研究的结果Eur J Med Res. 1998; 3(11):511- 14I.
Ammon JP. Modulation of the immune system by Boswellia serrata extracts and boswellic acids. 齿叶乳香提取物和乳香酸对免疫系统的调节 Phytomedecine. 2010; 17(11):862-7.
Mattmiller SA et al.Regulation of inflammation by selenium and selenoproteins: impact on eicosanoid biosynthesis. 硒和硒蛋白对炎症的调节:对类花生酸生物合成的影响。 J Nutr Sci. 2013; 2:e28.
Bonaventura P et al. Zinc and its role in immunity and inflammation. 锌及其在免疫和炎症中的作用 Autoimmun Rev. 2015; 14(4):277-85.
Prasad AS. Zinc is an antioxidant and anti-inflammatory agent: its role in human health. 锌作为抗氧化剂和消炎剂在人类健康中的作用 Front Nutr. 2014.
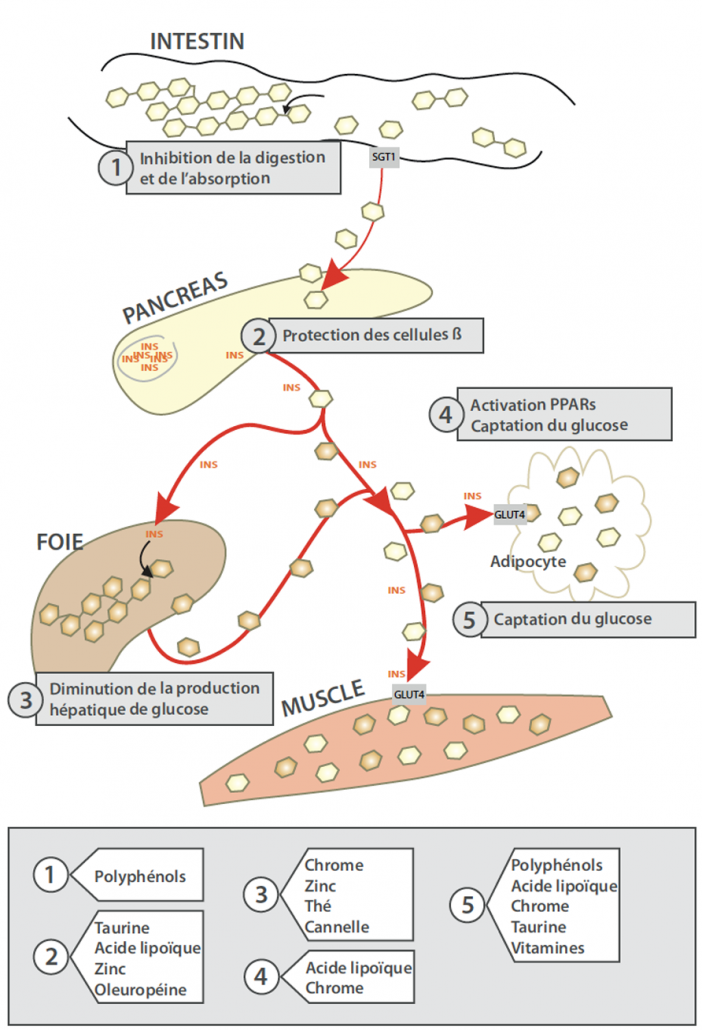
图5:微营养素对碳水化合物稳态的作用方式 (selon Hanhineva K et al. Int J Mol Sci 2010; 11(4):1365-1402)
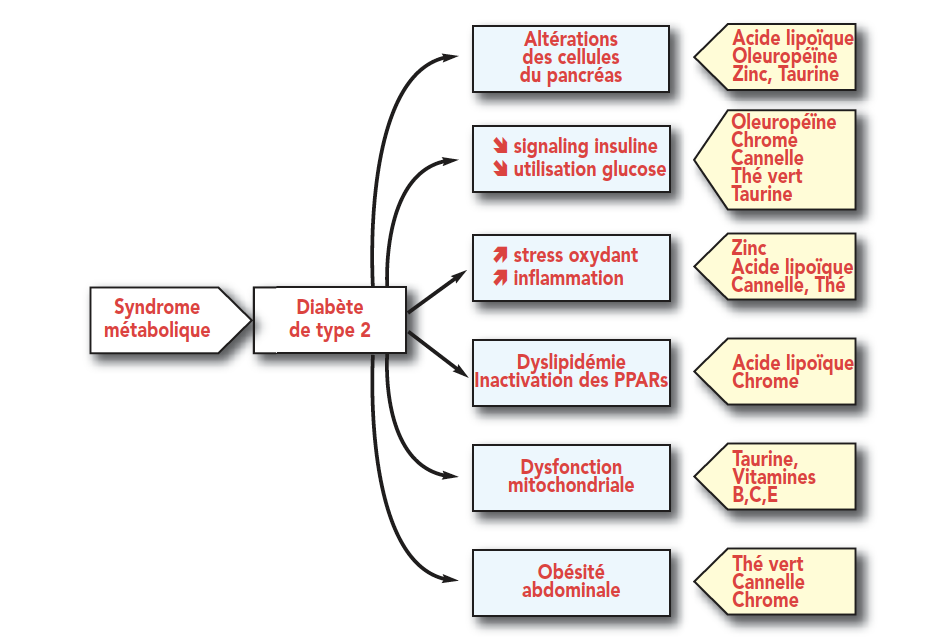
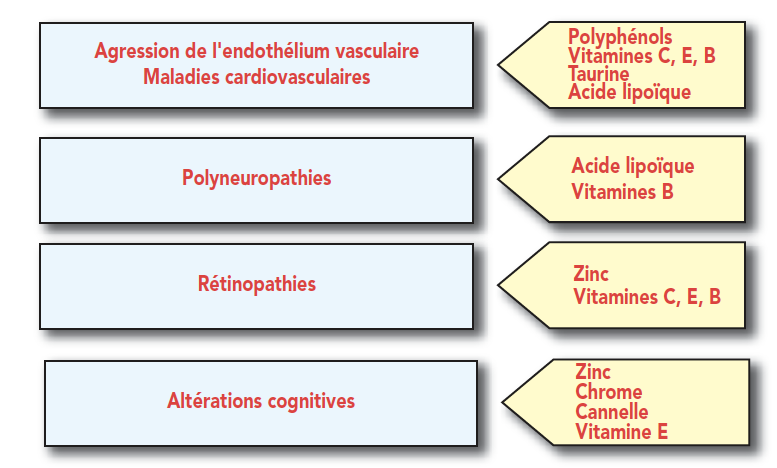
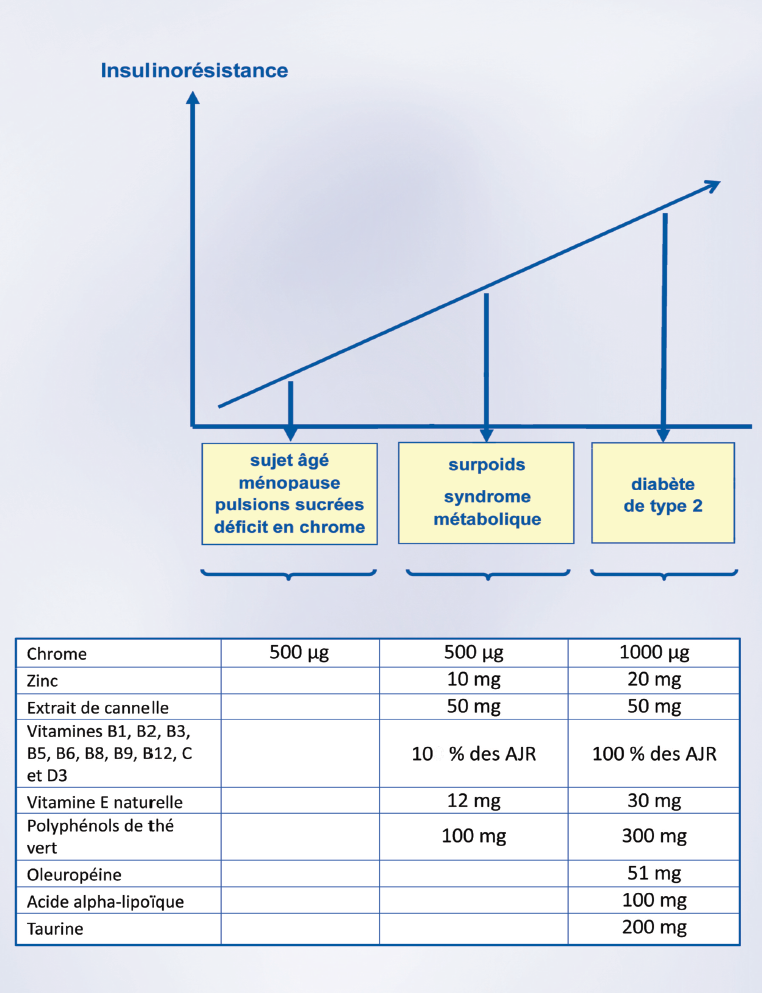
图6:微量营养素,代谢综合症和2型糖尿病
图7:2型糖尿病的微营养素和并发症
图8:对胰岛素抵抗具有活性的微量营养素:汇总表。
参考文献
Grundy SM. Obesity, metabolic syndrome and cardiovascular disease. 肥胖、代谢综合症和心血管疾病J Clin Endocrinol Metab, 2004; 89: 2595-2600.
Carey DG et al. Abdominal fat and insulin resistance in normal and overweight women: direct measurements reveal a strong relationship in subjects at both low and high risk of 正常和超重妇女的腹部脂肪和胰岛素抵抗:直接测量显示,在低和高NIDDM风险受试者中都有密切关系 Diabetes, 1996;45:633-638.
Grimaldi A. Traité de diabétologie, 糖尿病的特质 2eme édition. Ed Médecine-Sciences, Flammarion, Janvier
Minich DM et Bland JS. Dietary management of the metabolic syndrome beyond 除了大量营养素外,代谢综合症的饮食管理 Nutr Rev. 2008; 66(8):429-44.
Bartlett HE, Eperjesi Nutritional supplementation for type 2 diabetes: A systematic review. 型糖尿病的营养补充:系统评价 Ophthalmic Physiol Opt, 2008, 28:503-523.
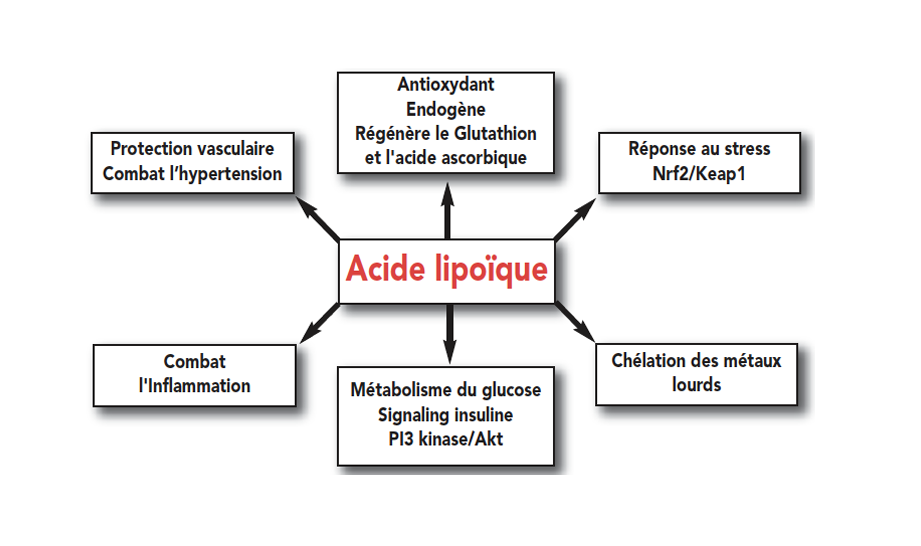
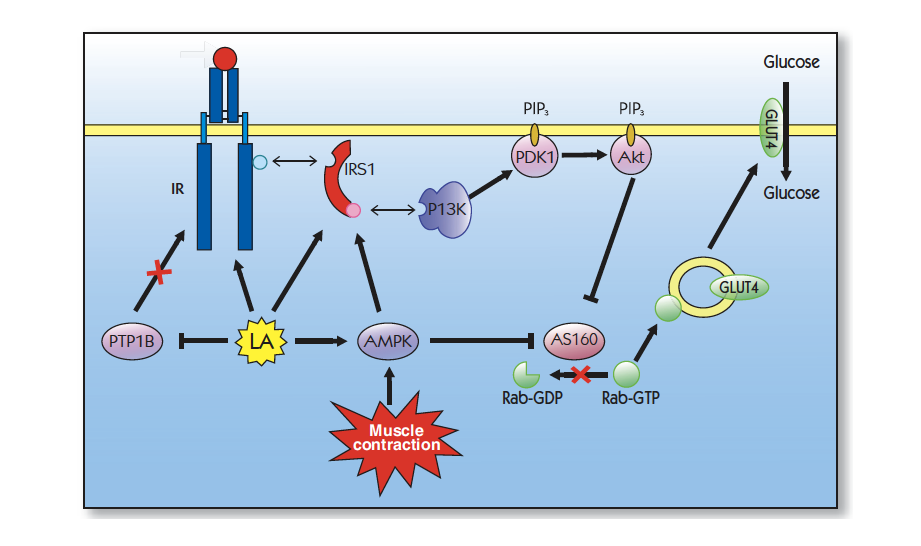
Shay et al. Alpha-lipoic acid as a dietary supplement: molecular mechanisms and therapeutical 硫辛酸作为膳食补充剂:分子机制和治疗潜力 Biochem Biophys Acta, 2009; 1790 (10):1149-1160.
Kamenova Improvement of insulin sensitivity in patients with type2 diabetes mellitus after oral administration of alpha lipoic acid. 口服α硫辛酸后改善2型糖尿病患者的胰岛素敏感性 Hormones 2006, 5(4):251-258.
Singh U, Jialal I. Alpha-lipoic acid supplementation and α硫辛酸补充剂和糖尿病 Nutr Rev, 2008, 66:646-657.
Golbidi S et al. Diabetes and Alpha lipoic acid. 糖尿病和α硫辛酸 Front 2011; 2:69
Manning PJ et The effect of lipoic acid and vitamin E therapies in individuals with the metabolic syndrome. 硫辛酸和维生素E治疗对代谢综合征患者的影响Nutr Metab Cardiovasc Dis. 2012, Mars.
Mcllduff CE, Rutkove SB.Critical appraisal of the use of alphalipoic acid in the treatment of symptomatic diabetic 评估使用α硫辛酸治疗有症状的糖尿病性多发性神经病 Ther Clin Risk Manag. 2011;7:377-385.
Tang J et Alpha-lipoic acid may improve symptomatic diabetic polyneuropathy. α硫辛酸可改善症状性糖尿病多发性神经病 Neurologist. 2007; 13:164-167.
Tankova T et al. Alpha-lipoic acid in the treatment of autonomic diabetic neuropathy (controlled, randomized, open-labelled study); α-硫辛酸治疗植物性糖尿病神经病变(对照随机开放标签研究)Rom J Intern Med. 2004;42:457-464.
Ziegler D et Efficacy and safety of antioxidant treatment with alpha-lipoic acid over 4 years in diabetic polyneuropathy: the NATHAN 1trial. 在糖尿病多发性神经病中使用α-硫辛酸抗氧化剂治疗4年以上的有效性和安全性:NATHAN 1试验。Diabetes Care, 2011;34(9):2054-60
Jacob S et Thioctic acid-Effects on insulin sensitivity and glucose metabolism. 硫辛酸对胰岛素敏感性和葡萄糖代谢的影响 Biofactors.1999; 10: 169-174.
Evans JL et al. Pharmacokinetics, tolerability and fructosamine-lowering effect of a novel, controlled-release formulation of alpha-lipoic acid. 新型α-硫辛酸控释制剂的药代动力学、耐受性和果糖胺降低作用Endocr 2002;8:29-35.
Konrad T et Alpha lipoic treatment decreases serum lactate and pyruvate concentrations and improves glucose effectiveness in lean and obese patients with type 2 diabetes. α硫辛酸治疗可降低2型糖尿病的肥胖和肥胖患者的血清乳酸和丙酮酸浓度,并改善葡萄糖有效性 Diabetes care.1999; 22:280-287.
Heitzer T et Beneficial effects of alpha-lipoic acid and ascorbic acid on endothelium-dependent, nitric oxide- mediated vasodilation in diabetic patients: relation to parameters of oxidative stress. α-硫辛酸和抗坏血酸对糖尿病患者内皮依赖性一氧化氮介导的血管舒张的有益作用:与氧化应激参数的关系 Free Radic Biol Med. 2001; 31:53-61
Haak E et al. Effects of alpha-lipoic acid on microcirculation in patients with peripheral diabetic α-硫辛酸对周围型糖尿病神经病患者微循环的影响 Exp Clin endocrinol diabetes.2000;108:168-174
Tankova T et Alpha-lipoic acid in the treatment of autonomic diabetic neuropathy (controlled, randomized, open-labelled study) α-硫辛酸治疗植物性糖尿病神经病变(对照随机开放标签研究); Rom J Intern Med. 2004; 42:457-464.
Jin HY et The effect of alpha-lipoic acid on symptoms and skin blood flow in diabetic neuropathy. α-硫辛酸对糖尿病性神经病的症状和皮肤血流的影响 Diab Med.2007;24:1034-1038.
Tang J et Alpha-lipoic acid may improve symptomatic diabetic polyneuropathy. 硫辛酸可改善症状性糖尿病多发性神经病 Neurologist. 2007; 13:164-167.
Foster TS. Efficacy and safety of alpha-lipoic acid supplementation in the treatment of symptomatic diabetic α-硫辛酸补充剂治疗有症状的糖尿病性神经病的疗效和安全性Diabetes Educ. 2007;33:111-117.
Mijnhout GS et al. alpha lipoic acid for symptomatic peripheral neuropathy in patients with diabetes: a meta analysis of randomized controlled trials. α硫辛酸治疗糖尿病患者的症状性周围神经病:随机对照试验的荟萃分析Int J 2012; 2012:456279.Janv 26
Santora A et al. The effects of chronic taurine supplementation on motor learning. 长期补充牛磺酸对运动学习的影响 Adv exp Med 2013;775:177-85
Idrissi AE et Taurine regulation of blood pressure and vasoactivity. 牛磺酸调节血压和血管活性Adv Exp Med Biol. 2013;775:407-25.
Abebe W, Mozaffari Role of taurine in the vasculature: an overview of experimental and human studies. 牛磺酸在脉管系统中的作用:实验和人体研究概述Am J Cardiovasc Dis. 2011;1(3): 293-311.
Merheb M et Taurine intestinal absorption and renal excretion test in diabetic patients: a pilot study. 糖尿病患者牛磺酸肠吸收和肾排泄试验:一项初步研究 Diabetes Care, 2007;30: 2652-2654.
De Luca G et al. Taurine and osmoregulation: platelet taurine content, uptake and release in type 2 diabetic 牛磺酸和渗透调节:2型糖尿病患者的血小板牛磺酸含量,摄取和释放 Metabolism, 2001; 50:60-64.
De la Puerta C et Taurine and glucose metabolism. 牛磺酸和葡萄糖代谢 Nutr Hops. 2010; 25 (6) :910-9.
Ito T et The potential usefulness of taurine on diabetes mellitus and its complications. 牛磺酸对糖尿病及其并发症的潜在作用 Aminoacids 2012; 42(5) :1529-39.
Anuradha Curr Protein Pept Sci. 2009 ; 10(1) :8-17.
Xiao G et al. Oral taurine ameliorates NEFA-induced impairment in insulin sensitivity and beta cell function in obese and overweight non-diabetic 口服牛磺酸可改善肥胖和超重非糖尿病男性的NEFA诱导的胰岛素敏感性和β细胞功能损害 Diabetologia. 2008; 51 :139-146.
Kim SJ et al. Taurine-diabetes interaction: from involvement to protection. 牛磺酸与糖尿病的相互作用:从参与到保护J Biol Regul Homeost Agents. 2007;21 (3-4) :63-77.
Nandhini AT et al. Taurine modifies insulinsignalling enzymes in the fructose-fed insulin resistant rats 果糖喂养的胰岛素抵抗大鼠中牛磺酸修饰的胰岛素信号转导酶. Diabetes 2005; 31(4Pt1):337-44.
Hansen SH et J Biomed Sci. 2010 ; 17(Suppl1) :S23.
Schaffer SW et Role of antioxidant activity of taurine in diabetes. 牛磺酸抗氧化活性在糖尿病中的作用Can J Physiol Pharmacol. 2009; 87(2):91-9.
Galleano M et Flavonoids and the metabolic syndrome. 类黄酮与代谢综合征Ann NY Acad Sci, 2012, vol 1259, 87-94.
Hanhineva K et al. Impact of dietary polyphenols on carbohydrate metabolism; 膳食多酚对碳水化合物代谢的影响 Int J Mol Sci; 2010; 11(4):1365- 1402.
Wainstein J et Olive leaf extract as a hypoglycaemic agent in both human diabetic subjects and in rats. 橄榄叶提取物在人类糖尿病患者和大鼠中均作为降血糖药 J Med Food. 2012; 15(7): 605-10.
De Bock M et al. Olive (Olea Europea) leaf polyphenols improve insulin sensitivity in middle-aged overweight men: a randomized, placebo-controlled, crossover 橄榄叶(Olea Europea)叶多酚可改善中年超重男性的胰岛素敏感性:一项随机安慰剂对照交叉试验 PLosOne, 2013; 8 (3).
Qin B, Anderson RA. Cinnamon: potential role in the prevention of insulin resistance, metabolic syndrome and type 2 肉桂:预防胰岛素抵抗,代谢综合征和2型糖尿病的潜在作用 J Diabetes Sci Technol. 2010; 4(3):685-93.
Jiao L et Proanthocyanidins are the major anti-diabetic components of cinnamon water extract. 原花青素是肉桂水提取物的主要抗糖尿病成分 Food Chem Toxicol. 2013; 56:398-405.
Anderson RA et al. Isolation and characterization of polyphenols type-A polymers from cinnamon with insulin- like biological 具有胰岛素样生物活性的肉桂中多酚A型聚合物的分离和表征 J Agric Food Chem. 2004, 52 (1):65- 70.
Magistrelli A, Chezem Effect of ground cinnamon on postprandial blood glucose concentration in normal-weight and obese adults. 肉桂粉对正常体重和肥胖成年人餐后血糖浓度的影响J Acad Nutr Diet. 2012; 112(11):1806-9.
Akilen R et al. Effect of short-term administration of cinnamon on blood pressure in patients with prediabetes and type 2 短期服用肉桂对糖尿病前期和2型糖尿病患者血压的影响 Nutrition, 2013; Jul 15.
Lu T et al. Cinnamon extract improves fasting blood glucose and glycosylated haemoglobin level in Chinese patients with type 2 肉桂提取物改善中国2型糖尿病患者的空腹血糖和糖基化血红蛋白水平 Nutr Res. 2012;32 (6):408-12
Ranasinghe P et Efficacity and safety of true cinnamon as a pharmaceutical agent in diabetes: a systematic review and meta analysis.肉桂作为糖尿病药物的功效和安全性:系统评价和荟萃分析Diab Med. 2012; 29(12):1480-92.
Akilen R et Cinnamon in glycaemic control: Systematic review and meta analysis. 肉桂在血糖控制中的应用:系统评价和荟萃分析Clin Nutr 2012; 31(5):609-15.
Davis PA, Yokoyama Cinnamon intake lowers fasting blood glucose: meta-analysis. 肉桂摄入量降低空腹血糖:荟萃分析 J Med Food. 2011; 14(9):884- 9.
Polychronopoulos E et Effects of black and green tea consumption on blood glucose levels in non obese elderly men and women from Mediterranean islands (MEDIS study). 食用红茶和绿茶对地中海岛屿非肥胖老年男女血糖水平的影响(MEDIS研究) Eur J Nutr. 2008; 47110-6.
Mackenzie T et The effect of an extract of green and black tea on glucose control in adults with type2 diabetes mellitus: double- blind randomized study. 绿茶和红茶提取物对2型糖尿病成年人血糖控制的影响:双盲随机研究 Metabolism. 2007;56 (10):1340-4.
Herder C and the InterAct Tea consumption and incidence of type 2 diabetes in Europe: the EPIC-Inter Act Case-Cohort study. 欧洲的茶摄入量和2型糖尿病的发病率:EPIC-Inter Act案例研究 PloS One 2012; 2 (5):236910.
Cao H et Green tea polyphenol extracts regulates the expression of genes involved in glucose uptake and insulin signalling in rats fed a high fructose diet. 绿茶多酚提取物调节高果糖饮食大鼠的葡萄糖摄取和胰岛素信号传导相关基因的表达 J Agric Food Chem. 2007a, 55:6372-378.
Fukino Y et Randomized controlled trial for an effect of green tea-extract powder supplementation on glucose abnormalities. 绿茶提取物补充粉对葡萄糖异常影响的随机对照试验 Eur J Clin Nutr. 2007 Jun.
Wiernsperger N, Rapin J. Trace elements in gluco metabolic disorders: an update. 糖代谢紊乱中的微量元素:更新 Diabetol Metab Syndr. 2010; 19:2:70.
Bao W et al. Dietary iron intake, body iron stores and the risk of type 2 diabetes: A systematic review and meta-analysis. 膳食铁摄入、体内铁储备和2型糖尿病的风险:系统评价和荟萃分析 BMC Med. 2012; 10:119.
Abdollahi M et al. Effect of chromium on glucose and lipid profiles in patients with type 2 diabetes: a meta-analysis review of randomized trials. 铬对2型糖尿病患者血糖和血脂谱的影响:一项随机试验的荟萃分析J Pharm Sci. 2013; 16(1):99-114.
Chen YL et al. The effect of chromium on inflammatory markers, 1st and 2nd phase insulin secretion in type 2 diabetes. 铬对2型糖尿病患者炎症标志物,一期和二期胰岛素分泌的影响 Eur J Nutr. 2013; Mars 14.
Sharma S et al. Beneficial effects of chromium supplementation on glucose, HbA1C and lipid variables in individuals with newly onset type-2 diabetes. 铬补充对新发2型糖尿病患者葡萄糖,HbA1C和脂质变量的有益作用
Jain SK et al. Effect of chromium supplementation on circulating levels of insulin, TNFa, oxidative stress, and insulin resistance in type 2 diabetic subjects: a randomized, double- blind, placebo-controlled study. 补充铬对2型糖尿病受试者体内循环血胰岛素,TNFα,氧化应激和胰岛素抵抗的影响:一项随机双盲安慰剂对照研究Mol Nutr Food Res. 2012; 56(8):1333-41.
Lai MH. Antioxidant effect and insulin resistance improvement of chromium combined with vitamin C and E supplementation for type 2 diabetes mellitus. 铬联合维生素C和E补充2型糖尿病的抗氧化作用和胰岛素抵抗的改善 J Clin Biochem Nutr. 2008; 43 (3):191-8.
Albarracin CA et al. Chromium picolinate and biotin combination improves glucose metabolism in treated, uncontrolled overweight to obese patients with diabetes type2. 吡啶甲酸铬和生物素的结合改善了肥胖的2型糖尿病患者的糖代谢,这种糖的治疗是不受控制的超重 Diabetes Metab Res rev. 2008; 24 (1):41-51.
Hua Y et al. Molecular mechanisms of chromium in alleviating insulin resistance; 铬减轻胰岛素抵抗的分子机制 J Nutr Biochem. 2012; 23(4):313-9.
Anton SD et al. Effects of chromium picolinate on food intake and satiety. 吡啶甲酸铬对食物摄入和饱腹感的影响 Diabetes Technol Ther. 2008; 10 (5):405- 412.
Vardatsikos G et al. Insulino-mimetic and antidiabetic effects of zinc. 锌的模拟胰岛素和抗糖尿病作用 J Inorg Biochem. 2013; 120: 8-17.
Miao X et al. Zinc homeostasis in the metabolic syndrome and diabetes. 代谢综合症和糖尿病中的锌稳态Front Med.2013; 7 (1):31-52.
Capdor J et al. Zinc and glycemic control: a meta-analysis of randomised placebo controlled supplementation trials in humans. 锌和血糖控制:人类随机安慰剂对照补充试验的荟萃分析 J Trace Elem Med Biol. 2013; 27(2):137- 42.
Jayawardena R et al. Effects of zinc supplementation on diabetes mellitus: a systematic review and meta-analysis. 补锌对糖尿病的影响:系统评价和荟萃分析 Diabetol Metab Syndr. 2012; 4(1):13.
Mezza T et al. Vitamin D deficiency: a new risk for type 2 diabetes. 维生素D缺乏症:2型糖尿病的新危险 Ann Nutr Metab. 2012,61(4) :337-4.
Kristakoss G et al. Vitamin D : beyond bone. 维生素D:不止是骨骼 Ann NY Acad Sci,2013 ;1287 :45-58.
We may request cookies to be set on your device. We use cookies to let us know when you visit our websites, how you interact with us, to enrich your user experience, and to customize your relationship with our website.
Click on the different category headings to find out more. You can also change some of your preferences. Note that blocking some types of cookies may impact your experience on our websites and the services we are able to offer.
Essential Website Cookies
These cookies are strictly necessary to provide you with services available through our website and to use some of its features.
Because these cookies are strictly necessary to deliver the website, you cannot refuse them without impacting how our site functions. You can block or delete them by changing your browser settings and force blocking all cookies on this website.
Other external services
We also use different external services like Google Webfonts, Google Maps and external Video providers. Since these providers may collect personal data like your IP address we allow you to block them here. Please be aware that this might heavily reduce the functionality and appearance of our site. Changes will take effect once you reload the page.
Google Webfont Settings:
Google Map Settings:
Vimeo and Youtube video embeds:
Privacy Policy
You can read about our cookies and privacy settings in detail on our Privacy Policy Page.